
Translate this page into:
Strategies & recent development of transmission-blocking vaccines against Plasmodium falciparum
Reprint requests: Dr Neeru Singh, National Institute for Research in Tribal Health (NIRTH), [Formerly Regional Medical Research Centre for Tribals (RMRCT)], NIRTH (ICMR) campus, Nagpur Road, Garha, Jabalpur 482 003, Madhya Pradesh, India e-mail: neeru.singh@gmail.com
-
Received: ,
This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under the identical terms.
This article was originally published by Medknow Publications & Media Pvt Ltd and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Transmission blocking malaria vaccines are aimed to block the development and maturity of sexual stages of parasite within mosquitoes. The vaccine candidate antigens (Pfs25, Pfs48/45, Pfs230) that have shown transmission blocking immunity in model systems are in different stages of development. These antigens are immunogenic with limited genetic diversity. Pfs25 is a leading candidate and currently in phase I clinical trial. Efforts are now focused on the cost-effective production of potent antigens using safe adjuvants and optimization of vaccine delivery system that are capable of inducing strong immune responses. This review addresses the potential usefulness, development strategies, challenges, clinical trials and current status of Plasmodium falciparum sexual stage malaria vaccine candidate antigens for the development of transmission-blocking vaccines.
Keywords
Malaria
Plasmodium falciparum
sexual stage antigens
transmission blocking vaccine

Malaria transmission blocking vaccine at a glance
Malaria is considered as a major global health problem, with an estimated 214 million cases and 438,000 malaria-related deaths worldwide in 20151. Plasmodium falciparum is responsible for a majority of malaria cases in humans. The emergence of insecticide-resistant mosquitoes and increase in parasite resistance to antimalarial drugs enhanced the need for effective vaccine development2. Multiple stages (pre-erythrocytic, erythrocytic and sexual stage) of the life cycle of malaria parasite are being targeted for vaccine development. Transmission blocking vaccines (TBVs) are focused against sexual stages or sporogonic-specific antigens. These are designed to block the development of sporogonic stages of parasite inside the mosquito thereby reducing mosquito infectivity and prohibiting the spread of the disease3. The target antigens for TBVs are divided into two groups, namely, pre-fertilization and post-fertilization antigens. Pre-fertilization antigens are expressed on the surface of gametocytes and gametes of malaria parasites, such as Pfs48/45, Pfs47 and Pfs2304. These proteins belong to a family that contains six-cysteine domains5. Pfs230 and Pfs48/45 are major gamete surface antigens that induce antibody responses in naturally exposed individuals67 and are associated with transmission reducing immunity8. Pfs25 is a post-fertilization antigen expressed on the surface of zygote and ookinete and has shown strong immunogenicity with limited antigenic polymorphism910. The antibodies that only target conformational epitopes of these proteins depend on the proper folding of cysteine-rich proteins and exact formation of disulphide bridges1112. These sexual stage antigens induce antibodies in the human host that interfere with the parasite development. Thus, transmission blocking takes place inside the mosquito vector and is antibody mediated13. In vitro studies of the transmission-reducing immune response in animal models1415 have shown a significant reduction in parasite development, which has led to the development of TBVs as part of malaria control and elimination strategy.
Expression of TBV target proteins in P. falciparum
More than hundred genes are expressed in the parasite life cycle, however, only a few of them have been cloned and studied for vaccine candidate. A unique Plasmodium gene superfamily encoding proteins that share six-cysteine domains are expressed during the sexual stages. The Pfs48/45 family has 12 distinct members namely, Pfs230, Pfs48/45, Pfs230p, Pfs47, P52, P36, Pf41, Pf38, Pf12, P12p, Pf92 and sequestrin16. Among these proteins, Pfs230, Pfs48/45 and Pfs47 play a critical role in the parasite development17. The six-cysteine family is conserved throughout all Plasmodium species and characterized by partially conserved cysteine-rich double domains having approximately 350 amino acids in length contributing to the tertiary structure of the proteins. Most of these proteins are localized on the parasite surface and some of these are known to play a role in cell-cell interaction518. The immunogenic proteins (Pfs48/45, Pfs47, Pfs230, and Pfs25) are important for fertilization process and other vital functions of parasite life cycles13. These antigens have the ability to boost the immune system either by vaccination or naturally during the infection19. These specific characteristics of the sexual stage antigens make the proteins interesting for study the development biology of the parasite in the mosquito vector and ultimately the possible vaccine targets (Tables I & II).
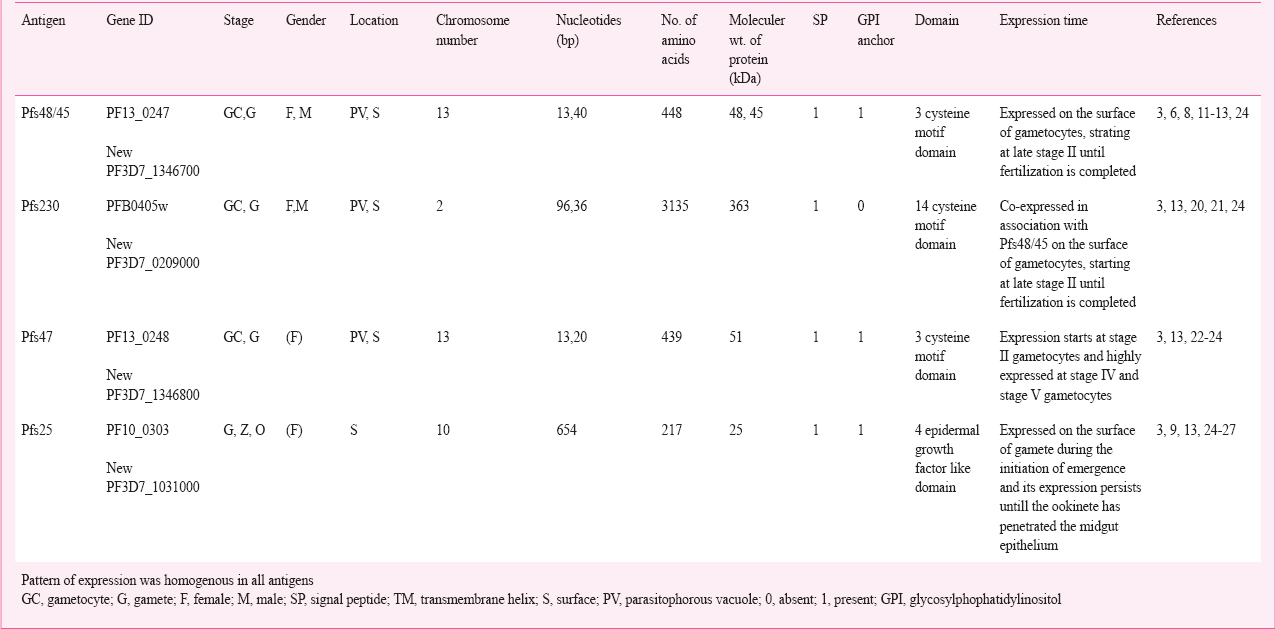
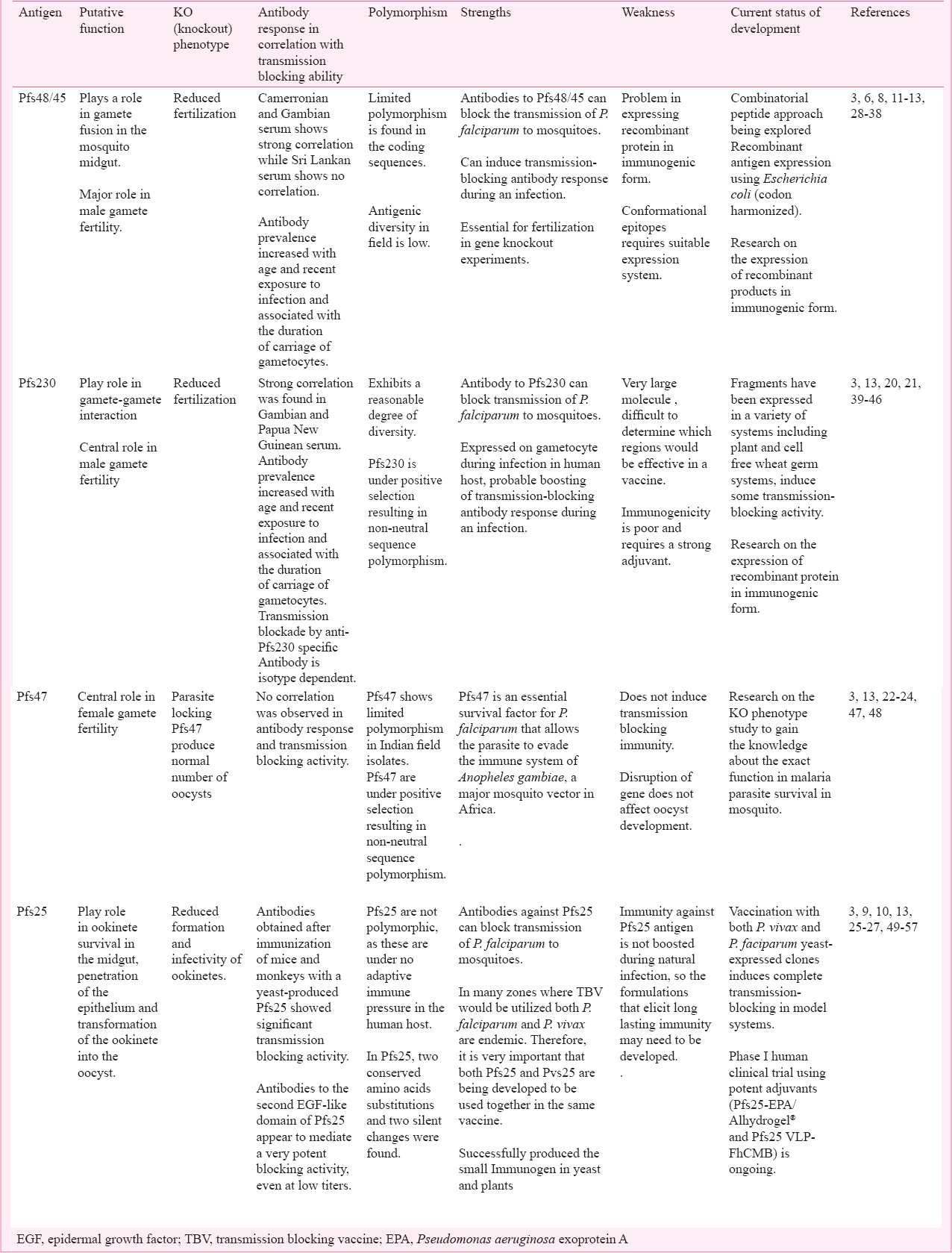
Potential of TBV candidates
Pfs48/45: The Pfs48/45 antigen plays a role in the male/female gamete fusion in the mosquito midgut5. Disruption of the Pfs48/45 gene in P. falciparum and P. berghei has demonstrated a central role in male gamete fertility13. Fertilization and zygote formation are strongly reduced in Pfs48/45 knockout parasites, but production of gametocyte and differentiation into gametes remains unaffected17. The Pfs48/45 antigen induces antibody responses in naturally exposed individuals which are associated with functional transmission-reducing immunity68. Transmission-blocking monoclonal antibodies that recognize B-cells epitopes on Pfs48/45 seem to block fertilization with the presence of complement proteins11 as well as without complement12. The ability to stimulate the antibody response upon encounter with the natural infection as seen in the field makes the exceptionally valuable capability of vaccine boosting in the endemic areas. Data from hyperendemic Papua New Guinea (PNG) show that seroprevalence increases with age, suggesting that anti-Pfs48/45 response develops immunological memory28. Increasing antibody titres against Pfs48/45 have also been observed with recent exposure to malaria infection in PNG29. Study from Gambian and Cameroonian populations showed strong correlation between antibody response and transmission reducing activity against Pfs48/45 antigen3031, while anti-Pfs48/45 response in serum of Sri Lankan population did not show any correlation32. The antibody response against Pfs48/45 is enhanced by simultaneous exposure of gametocyte and is also related to the extent of gametocyte carriage in Tanzania33. Conversely, the studies carried out in Senegal and Cameroon conclude that transmission blocking immunity depends on age, antibody titres, episode of malaria infections and duration of gametocyte carriage34. Genetic polymorphism is a major problem in malaria vaccine development. The Pfs48/45 gene is less polymorphic in comparison to other erythrocytic and pre-erythrocytic stage specific antigens353637 with a minimum number of amino acid substitutions38. These observations suggest that this transmission blocking antigen is conserved at the protein level, making it a good candidate for multistage, multivalent vaccine. Fusion of Pfs48/45 to glutamate rich protein (GLURP) antigen is one of the recent approaches towards the development of multi-stage malaria vaccine which can target both transmission and asexual parasite life cycle stages39. All these findings show that Pfs48/45 is an important candidate antigen for the development of transmission blocking vaccine.
Pfs230: Pfs230 is a 363 kDa (3135 amino acid) protein and a potent antigen of malaria transmission blocking vaccine. It is involved in the fertilization of macrogametocytes by microgametocytes20. Male gamete with a disrupted Pfs230 gene is incapable of interacting with erythrocytes and unable to form exflagellation center20. Evidence from a mutant analysis study has shown that in the absence of Pfs48/45, the Pfs230 protein is not retained on the surface of gametes indicating that tethering of Pfs230 is mediated by Pfs48/4521. Transmission reducing activity of Pfs230 is isotype dependent40 and blocks gamete formation via complement-mediated lysis41. Radiolabelled antibodies against Pfs230 are able to bind to the surface of gametes and reduce the P. falciparum infectivity to mosquitoes7. Antibody responses against the Pfs230 were observed in naturally exposed individuals4243. The strong correlation between transmission blocking activity and anti-Pfs230 antibody response was found in Gambian30 and Papua New Guinean populations44. The antibody response in Cameroonian populations showed weak correlation45 while no correlation was observed in Sri Lankan populations32. A study from Tanzania showed that the antibody level against Pfs230 increased with age, recent exposure to infection and associated with the gametocytes carriage33. Pfs230 exhibits a reasonable degree of diversity46. These all characteristics of Pfs230 make this antigen an important promising candidate for TBVs.
Pfs47: The Pfs47 antigen is a contiguous paralog of Pfs48/45 located on 1.5Kb apart from Pfs48/45 and arranged tandemly on chromosome number 1322. Protein expression of Pfs47 is sex-specific and expressed on the surface of female gametocyte and gamete2324. Study on Pfs47 demonstrates that it is not crucial for female fertility47. Parasite lacking Pfs47 through targeted gene disruption produced a normal number of oocysts and anti-Pfs47 monoclonal antibodies were unable to inhibit oocysts development47. These characteristics reduced the potential of Pfs47 as a good TBV target. Pfs47 inhibits the Jun-N-terminal kinase (JNK) mediated apoptosis process by inhibiting the activation of caspases; this inhibition leads to inadequate nitration reaction and thus parasite becomes invisible to complement-like system in the mosquito midgut48. Interruption of immunomodulatory action of Pfs47 gene inside the mosquito vector may turn out to be a convincing methodology to decrease malaria transmission47. A study carried out at National Institute for Research in Tribal Health (NIRTH), Jabalpur on serum samples of Indian patients showed that seroprevalence against Pfs47 antigen was highest among other TBV candidates (i.e. Pfs48/45 and Ps230). Pfs47 also showed limited genetic polymorphism in the Indian field isolates (Chaturvedi et al, unpublished observations). These findings support the candidacy of Pfs47 as a potential target of transmission blocking vaccine. However, more studies are needed to gain the knowledge of immunomodulatory activity of Pfs47 in other mosquito vectors.
Pfs25: Pfs25 is considered as one of the most important transmission blocking vaccine candidate antigens, expressed on the surface of zygote and ookinetes. Gene knockout experiments suggest that this protein is important for the parasite to survive inside the mosquito midgut25. Further, double-knockout study on P. berghei shows that the loss of P25 antigen reduces the ookinete invasion into the midgut epithelial cells2627. The Pfs25 protein is expressed only in the mosquito host and antibody raised against the recombinant Pfs25 protein stops the parasite development within the mosquito vector49. The significance of post-fertilization antigen is long lasting immunogenicity and less antigenic variations91050. An immunogenic form of Pfs25 expressed in yeast51 shows effective transmission blocking activity in membrane feeding assays52. To enhance the efficacy of antibody response, the chimeric form of Pfs 25-28 was expressed in yeast and tested on mice. Pfs25-28 showed the more potent response to block the oocyst formation as compared to individual proteins i.e. either Pfs-25 or Pfs-28 alone53. Intramuscular administration of Pfs25 in mice elicited the potential transmission-blocking antibodies that resulted in more than 90 per cent reduction in oocyst numbers in the mosquito midgut54. Phase I clinical trial of modified forms of Pfs25 called TBV25H (consisting of extra his-tag at the C- terminus of protein) in human volunteers with aluminum hydroxide adjuvant showed 50 per cent reduction in P. falciparum infectivity51. The TBV25H constructs utilized as a part of the initial tests and human trials shows antigenicity nearly to the native molecules51. Recombinant Pfs25 expressed in yeast linked to the outer membrane protein complex (OMPC) of Neisseria meningitidis serogroup B with aluminium hydroxyphosphate formulations has shown stronger anti-Pfs25 antibody response than Pfs25 alone with montanide ISA 720 at the same measurements55. Recombinant Pfs25H in conjugation with exoprotein A (EPA) of Pseudomonas aeruginosa has been produced and used to make a cGMP pilot lot to use in Phase I human clinical trials in the United States56. The practical assessment of Pfs25 DNA vaccine done by in vivo electroporation in olive baboons showed potent immunogenicity against malaria57. Consequences of DNA-based immunization in non-human primates give the premise to further assessment in human volunteers. Endeavours are presently centered on the clinical advancement of Pfs25-CP VLP, containing Pfs25 fused to the alfalfa mosaic virus coat protein (CP) expressed in Nicotiana benthamiana plants using a tobacco mosaic virus (TMV) based “launch vector” technology. Administration of one or two doses of Pfs25-CP VLPs with adjuvant Alhydrogel® in mice stimulates the antibodies that have shown absolute transmission blocking activity58. A clinical trial with this immunogenic preparation is ongoing in the United States (https://clinicaltrials.gov/ct2/show/NCT02013687). Trials with clinical grade formulations of Pfs25 molecules involves the preparation of different constructs in immunogenic forms and the testing of safe adjuvant59.
Other important transmission-blocking vaccine candidate antigens: Intensive research on TBVs has discovered several other potential vaccine candidates that include other surface antigens (gamete, zygote, ookinete) and a few important mosquito proteins that are required by the parasite for its maturation in a vector. Pfs28 is expressed on the ookinete surface as an antigen distinct from Pfs25 and plays a role in ookinete protection from stomach enzymes6061. Knockout phenotypes of Pfs28 significantly reduced mosquito infectivity26. Potent Pfs28 was expressed as a chimeric protein in combination with Pfs25 to enhance the transmission blocking activity53. Vector molecules such as carboxypeptidase B (CPB) and aminopeptidase N (APN) play an important role in the development of P. falciparum inside the mosquito and, therefore, are considered as TBV candidates62636465. These candidates are conserved and elicit transmission reducing antibodies6667. However, efforts are underway that attempt to understand the mode of action and production of these antigenic targets in immunogenic forms. Other important transmission blocking vaccine candidate genes that are currently in the development stage6869 are discussed in the Table III.
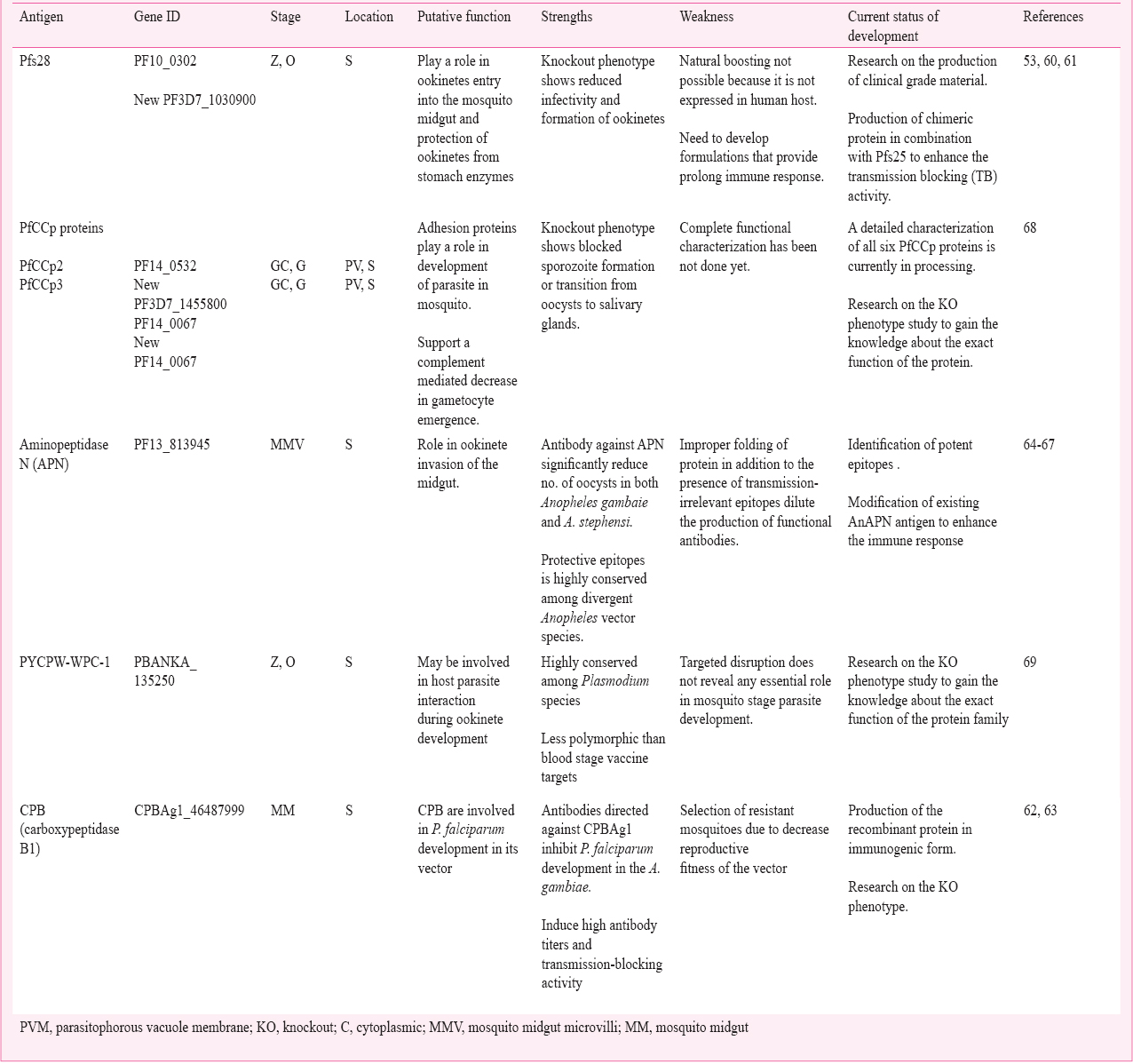
Production of TBVs in plant based system
The requirement for a new approach for expression of large quantities of vaccine antigen with a good safety profile is important. Plant-based expression systems may be an appropriate choice for the cost-effective production of TBVs. The plant-based expression system provides all the advantages of a eukaryotic expression system with high purity and stability. Various TBV candidates have been successfully produced in plant system707172 with high recovery and desired immunogenicity (Table IV). Algae are also a promising system for the production of TBV that are orally delivered to avoid expensive purification and injectible delivery737475. These results sustain the feasibility of expressing TBV antigens in the plant based system.

Effective adjuvants and vaccine delivery system
An efficient malaria vaccine against important developmental phases of the parasite life cycle, required to stimulate appropriate humoral and cellular immune responses. Transmission blocking immunity is governed by both cell-mediated and antibody-mediated effector mechanisms7677. However, the humoral immune response seems to be a key player in transmission blocking immunity as compared to cell-mediated response78. Presently authorized human immunization adjuvants like alum, may enhance the antibody generation, but are poor stimulators of cellular effector systems, while strong cell stimulants, such as Freund's adjuvant are found reactogenic for human utilization. Several methods of antigen presentation i.e. liposomes mediated delivery, emulsification in Freund's adjuvant and addition of bacterial protein are known to be safe and efficient strategies of vaccine delivery system that targets the different antigenic determinants to the host's immune system. Various studies have been done in analyzing the different adjuvant combination and effective delivery system to enhance the immunogenicity of vaccine candidates. Maltose binding protein fused with Pfs48/45 induces high antibody titres in mice and elicits functional antibody in standard feeding assays (SFA)15. This combination was found stable over a nine months period15. The intramuscular delivery system using novel carrier gel core liposomes encapsulated with Pfs25 showed significant and durable immune response79. To overcome the poor immunogenicity, recombinant Pfs25H conjugated to P. aeruginosa exoprotein A (EPA) has been developed and used in human clinical trials (https://clinicaltrials.gov/ct2/show/NCT01867463). Alhydrogel® adsorbed Pfs25-EPA nanoparticle vaccine significantly improved the Pfs25 antibody responses in mice56. In another study, non-classical concepts for vaccine delivery were found more suitable, in which vaccine delivery was done not only through parenteral, oral or mucosal routes but specifically via cutaneous immunization. Single inoculation and controlled release of antigen in mice, through biodegradable nano-microparticle technologies, can elicit long-lasting protective antibody titres with >85 per cent efficacy which remains effective for at least two years80.
Clinical trials
Presently, there are a small number of sexual stage vaccines under the preclinical advancement and Pfs25 is in Phase I clinical trial (https://clinicaltrials.gov/ct2/show/NCT01867463). Some clinical trials of Pfs25 became inactive due to systemic reactogenicity81 and unresolved issues with FDA (https://clinicaltrials.gov/ct2/show/NCT00977899). As the TBVs are intended to block the parasite development within the mosquitoes the vaccine efficiency should be measured at a community level via random field trials. Due to the knowledge gap between the trial sites and epidemiological data of different endemic regions, conducting the cluster randomized trials becomes challenging. Standard membrane feeding assays (SMFA) allows investigators to assess the capability of serum to decrease the mosquito infection1466. The data on the effect on transmission, immune responses in individuals and genetic diversity of sexual stage antigens are limited. Efforts are required to validate transmission inhibition and immune responses in humans living in endemic countries worldwide, as this knowledge will be important for vaccine development. Completed and ongoing clinical trials of sexual stage vaccines are discussed in Table V. Phase-I clinical trial of Pfs25 in a conjugate vaccine (Pfs25-EPA/Alhydrogel®) developed at National Institute of Allergy and Infectious Diseases (NIAID), USA was completed in healthy adults to assess the safety and immunogenicity with the collaboration of Programme for Appropriate Technology in Health-malaria Vaccine Initiative program PATH (MVI), NIAID and John Hopkins Bloomberg School of Public Health Centre for Immunization Research (CIR)82 (https://clinicaltrials.gov/ct2/show/NCT01434381). This vaccine was found increasingly immunogenic with each dose and induced transmission reducing antibody responses. Some safety issues were also reported in this clinical trial such as local and systemic adverse events82. Phase- I clinical trial of Pfs25 using montanide ISA-51 was also stopped due to safety issues. Following each vaccination, volunteers showed adverse events. Local and systemic adverse symptoms included swelling, tenderness, erythema, fever, nausea, headache, myalgia and arthralgia81. Pfs25 antigen expressed in plant system known as Fraunhofer's plant-derived malaria transmission-blocking vaccine, is in Phase- I clinical trial58 (https://clinicaltrials.gov/ct2/show/NCT02013687). Further human clinical trials are required to evaluate the vaccine candidate efficacy, health risk factors and cost-benefits assessments.
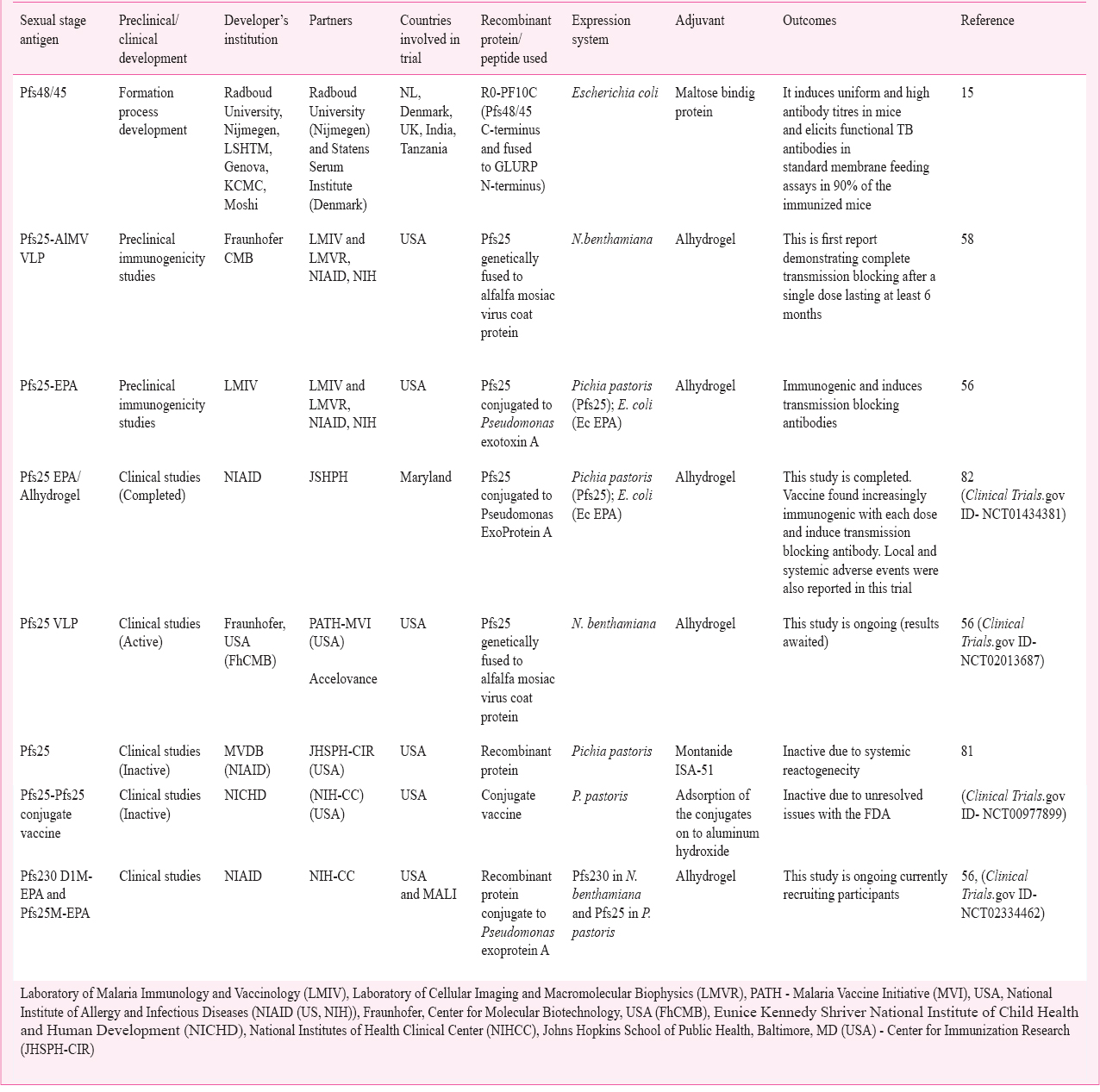
Challenges and opportunities
A new set of malaria vaccine candidates has entered into clinical trials and several new vaccine candidates are being developed. Various hurdles encountered in the development process of transmission blocking vaccine, includes (i) The problem of expressing TBV antigens in an appropriate conformation which is recognized by the antibody against the same epitopes; (ii) There are very limited known correlates of genetic diversity and immunity for transmission blocking antigens; (iii) Currently, only a few immune enhancing adjuvants are available, but these are very expensive; (iv) Analysis of transmission blocking activity needs more rigorously qualified assays and models to access the vaccine candidate efficacy; and (v) Funding constrant for TBV research and development.
The TBV development faces formidable challenges and collaborative approach is needed to solve scientific, economic and resource obstacles. Development of clinical trial sites in different regions with different epidemiological data should be done to analyze the safety and efficacy of vaccine candidates. A new advancement in this field can defeat these challenges. Advances in manufacturing industries can increase the large-scale production of vaccines, identification of potent adjuvants that can effectively boost the immune system, techniques like structural biology can predict the defensive epitopes, genomics and proteomic for novel antigen finding. Greater collaborations, new partnerships between research groups and policymaker's organization will be critical to developing a transmission blocking vaccine that could reduce the disease burden and transmission.
Concluding remarks
Malaria is a major global health problem and currently no vaccine is available to combat the disease. A promising transmission blocking vaccines is characterized and developed with clinical grade formulations during the last two decades. The completed clinical trials with Pfs25 formulations have been found immunogenic but shown some safety outcome issues like local and systemic adverse events8182. Further clinical trials are required in malaria endemic regions to assess the risk factors as well as the vaccine efficacy. Prefertilization antigens Pfs48/45 and Pfs230 have shown a strong association between sexual stage-specific antibody response and functional transmission reducing activity. Data from several studies showed that antibody response against Pfs230 and Pfs48/45 antigens was associated with the exposure and duration of gametocytes carriage33. Another important prefertilization antigen Pfs47 allows the parasite to evade the mosquito immune system by inhibiting the activation of apoptosis in mosquito midgut cells48. Beside all these characteristics, sexual stage antigens have also shown limited genetic polymorphism and strong potential for boosting the immune response424649. Further studies suggested that transmission blocking activity could likewise be obtained by targeting the mosquito components that are needed for the successful development of parasite inside the mosquito vector. These mosquito specific candidates include aminopeptidase N and carboxypeptidase B1, which are also able to induce an antibody response that significantly inhibits parasite development6365. However, further studies are needed to strengthen the candidacy of these antigens as a potent target of transmission blocking malaria vaccines.
Currently, many efforts for the development of transmission blocking malaria vaccine are focused on the production of TBV in plant-based expression system with a good safety profile, which are capable of inducing a strong immune response to reduce the malaria transmission717374. Completed clinical trials with Pfs25 formulations have been found immunogenic but shown some safety outcome issues like local and systemic adverse events8182. Human clinical trials are further required in malaria endemic regions to assess the risk factors and to evaluate the vaccine efficacy.
Future perspectives
Current worldwide approach for malaria control and elimination needs the vaccines that directly target the malaria transmission. For eradication, it is important that the vaccine provides potential contribution to reduce the infection rates, inhibits parasite development and thereby diminishing malaria mortality and morbidity by inhibiting the transmission. Malaria transmission blocking vaccines is a tool to reduce the mosquito infection and malarian transmission by inducing the immunity that breaks the cycle of malaria parasite between humans and mosquitoes.
Future efforts for the development of malaria transmission blocking vaccines for use as crucial components in malaria riddance must include (i) identification and functional characterization of those potent antigens and regulatory proteins which play crucial role in the development of the parasite in a mosquito vector; (ii) detection of molecular markers of sexual stage development; (iii) development of easily transferable in vitro culture system to better understand the dynamics between the multiplication of parasites, gametocyte biology and malaria transmission rates; (iv) improvement of an effective immunization method that would maintain the high antibody titre in the blood which will significantly affect parasite development in the mosquito; (v) identification of safe adjuvant combinations and dose optimization of vaccines; (vi) enhance the interest in the large scale production of malaria transmission blocking vaccines at the industrial level; (vii) clinical trials should be planned at the population level to check the safety and efficacy of the TBV; and (viii) development of multi-stage vaccine with the fusion of sexual stage antigen will also give future insight for malaria eradication.
Acknowledgment
The first author (NC) acknowledges the support of the Indian Council of Medical Research and Department of Biotechnology, Government of India for providing junior research fellowship. Authors appreciate the support and critical review given by Dr Altaf Lal, and Drs. Nirbhay Kumar and Aparup Das for their valuable comments and suggestion.
Conflicts of Interest: None.
References
- 2015. World Malaria Report 2015. Geneva: World Health Organization. Available from: http://www.who.int/malaria/publications/world-malaria-report-2015/report/en/
- 2010. World Malaria Report 2010. Geneva: World Health Organization. Available from: http://www.who.int/malaria/world_malaria_report_2010/en/
- Malaria transmission-blocking vaccine-how can their development be supported? Nature Med. 2000;6:241-4.
- [Google Scholar]
- Identification of additional members defines a Plasmodium falciparum gene superfamily which includes Pfs48/45 and Pfs230. Mol Biochem Parasitol. 1999;101:223-7.
- [Google Scholar]
- Sexual-stage antibody responses to P. falciparum in endemic populations. Curr Mol Med. 2006;6:223-9.
- [Google Scholar]
- Recombinant Pfs230, a Plasmodium falciparum gametocyte protein, induces antisera that reduce the infectivity of Plasmodium falciparum to mosquitoes. Mol Biochem Parasitol. 1995;75:33-45.
- [Google Scholar]
- Transmission-reducing immunity is inversely related to age in Plasmodium falciparum gametocyte carriers. Parasite Immunol. 2006;28:185-90.
- [Google Scholar]
- Plasmodium P25 and P28 surface proteins: potential transmission-blocking vaccines. Eukaryot Cell. 2007;6:1260-5.
- [Google Scholar]
- Minimal variation in a vaccine candidate from the sexual stage of Plasmodium falciparum. Mol Biochem Parasitol. 1989;32:101-3.
- [Google Scholar]
- Properties of epitopes of Pfs48/45, a target of transmission blocking monoclonal antibodies, on gametes of different isolates of Plasmodium falciparum. Parasite Immunol. 1990;12:587-603.
- [Google Scholar]
- Conserved and variant epitopes of target antigens of transmission-blocking antibodies among isolates of Plasmodium falciparum from Malaysia. Am J Trop Med Hyg. 1991;44:623-31.
- [Google Scholar]
- Proteins of the malaria parasite sexual stages: expression, function and potential for transmission blocking strategies. Parasitology. 2007;134:1911-29.
- [Google Scholar]
- The IC (50) of anti-Pfs25 antibody in membrane-feeding assay varies among species. Vaccine. 2010;17:4423-9.
- [Google Scholar]
- Correctly folded Pfs48/45 protein of Plasmodium falciparum elicits malaria transmission-blocking immunity in mice. Proc Natl Acad Sci USA. 2008;105:4301-5.
- [Google Scholar]
- Identification of protein complexes in detergent-resistant membranes of Plasmodium falciparum schizonts. Mol Biochem Parasitol. 2007;154:148-57.
- [Google Scholar]
- Gene expression in Plasmodium: from gametocytes to sporozoites. Int J Parasitol. 2004;34:1431-40.
- [Google Scholar]
- Discovery of gene function by expression profiling of the malaria parasite life cycle. Science. 2003;301:1503-8.
- [Google Scholar]
- Progress toward a transmission-blocking vaccine. In: Good MF, Saul J, eds. Molecular immunological consideration in malaria vaccine development. London: CRC Press, Boca Raton FL; 1994. p. :209-44.
- [Google Scholar]
- Malaria transmission-blocking antigen, Pfs230, mediates human red blood cell binding to exflagellating male parasites and oocyst production. Mol Microbio. 2006;61:991-8.
- [Google Scholar]
- Pfs230: from malaria transmission-blocking vaccine candidate toward function. Parasite Immunol. 2003;25:351-9.
- [Google Scholar]
- Pfs47, paralog of the male fertility factor Pfs48/45, is a female specific surface protein in Plasmodium falciparum. Mol Biochem Parasitol. 2006;149:216-22.
- [Google Scholar]
- The Plasmodium falciparum sexual development transcriptome: a microarray analysis using ontology-based pattern identification. Mol Biochem Parasitol. 2005;143:67-79.
- [Google Scholar]
- Sequential expression of antigens on sexual stages of Plasmodium falciparum accessible to transmission-blocking antibodies in the mosquito. J Exp Med. 1985;162:1460-76.
- [Google Scholar]
- Distinct roles for Pbs21 and Pbs25 in the in vitro ookinete to oocyst transformation of Plasmodium berghei. J Cell Sci. 2000;113:3419-26.
- [Google Scholar]
- P25 and P28 proteins of the malaria ookinete surface have multiple and partially redundant functions. EMBO J. 2001;20:3975-83.
- [Google Scholar]
- Do malaria ookinete surface proteins P25 and p28 mediate parasite entry into mosquito midgut epithelial cells? Malar J. 2005;4:15.
- [Google Scholar]
- Human antibody responses to epitopes on the Plasmodium falciparum gametocyte antigen Pfs 48/45 and their relationship to infectivity of gametocyte carriers. Am J Trop Med Hyg. 1992;46:711-9.
- [Google Scholar]
- Rapid onset of transmission-reducing antibodies in Javanese migrants exposed to malaria in Papua Indonesia. Am J Trop Med Hyg. 2006;74:425-31.
- [Google Scholar]
- Parasite infectivity and immunity to Plasmodium falciparum gametocytes in Gambian children. Parasite Immunol. 2004;26:159-65.
- [Google Scholar]
- Association between anti-Pfs48/45 reactivity and P. falciparum transmission-blocking activity in sera from Cameroon. Parasite Immunol. 1996;18:103-9.
- [Google Scholar]
- Plasmodium falciparum malaria transmission-blocking immunity under conditions of low endemicity as in Sri Lanka. Parasite Immunol. 1994;16:35-42.
- [Google Scholar]
- The dynamics of naturally acquired immune responses to Plasmodium falciparum sexual stage antigens Pfs230 & Pfs48/45 in a low endemic area in Tanzania. PLoS One. 2010;5:e14114.
- [Google Scholar]
- Plasmodium falciparum transmission blocking immunity in three areas with perennial or seasonal endemicity and different levels of transmission. Am J Trop Med Hyg. 2005;73:1090-5.
- [Google Scholar]
- Minimal variation in the transmission-blocking vaccine candidate Pfs48/45 of the human malaria parasite Plasmodium falciparum. Mol Biochem Parasitol. 1995;69:115-8.
- [Google Scholar]
- Genetic diversity in the block 2 region of the merozoite surface protein-1 of Plasmodium falciparum in central India. Malar J. 2012;11:78.
- [Google Scholar]
- Genetic variation in the Plasmodium falciparum circumsporozoite protein in India and its relevance to RTS, S malaria vaccine. PLoS One. 2012;7:e43430.
- [Google Scholar]
- Polymorphism in the gene encoding the Pfs48/45 antigen of Plasmodium falciparum. XI. Asembo Bay Cohort Project. Mol Biochem Parasitol. 2002;119:17-22.
- [Google Scholar]
- A multi-stage malaria vaccine candidate targeting both transmission and asexual parasite life-cycle stages. Vaccine. 2014;32:2623-30.
- [Google Scholar]
- Transmission blockade of Plasmodium falciparum malaria by anti-Pfs230-specific antibodies is isotype dependent. Infect Immunol. 1995;63:467-71.
- [Google Scholar]
- Complement-mediated lysis of Plasmodium falciparum gametes by malaria-immune human sera is associated with antibodies to the gamete surface antigen Pfs230. Infect Immun. 1997;65:3017-23.
- [Google Scholar]
- Naturally acquired immune responses to Plasmodium falciparum sexual stage antigens Pfs48/45 and Pfs230 in an area of seasonal transmission. Infect Immun. 2011;79:4957-64.
- [Google Scholar]
- Transmission blocking immunity to Plasmodium falciparum in malaria-immune individuals is associated with antibodies to the gamete surface protein Pfs230. Parasitology. 1999;119:425-33.
- [Google Scholar]
- Antibodies to Plasmodium falciparum gamete surface antigens in Papua New Guinea sera. Parasite Immunol. 1988;10:209-18.
- [Google Scholar]
- Reduction and enhancement of Plasmodium falciparum transmission by endemic human sera. Int J Parasitol. 2006;36:10-1.
- [Google Scholar]
- Limited polymorphism in Plasmodium falciparum sexual-stage antigen. Am J Trop Med Hyg. 2001;64:9-11.
- [Google Scholar]
- The human malaria parasite Pfs47 gene mediates evasion of the mosquito immune system. Science. 2013;340:1-3.
- [Google Scholar]
- Plasmodium falciparum evades mosquito immunity by disrupting JNKmediated apoptosis of invaded midgutcells. Proc Natl Acad Sci USA. 2015;112:1273-80.
- [Google Scholar]
- A region of Plasmodium falciparum antigen Pfs25 that is the target of highly potent transmission-blocking antibodies. Infect Immun. 2000;68:5530-8.
- [Google Scholar]
- Genetic polymorphism and natural selection in the malaria parasite Plasmodium falciparum. Genetics. 1998;149:189-202.
- [Google Scholar]
- Production, purification and immunogenicity of a malaria transmission-blocking vaccine candidate: TBV25H expressed in yeast and purified using nickel-NTA agarose. Biotechnology. 1994;12:494-9.
- [Google Scholar]
- Saccharomyces cerevisiae recombinant Pfs25 adsorbed to alum elicits antibodies that block transmission of Plasmodium falciparum. Infect Immun. 1994;62:5576-80.
- [Google Scholar]
- Saccharomyces cerevisiae secreted fusion proteins Pfs25 and Pfs28 elicit potent Plasmodium falciparum transmission-blocking antibodies in mice. Infect Immun. 1998;66:59-64.
- [Google Scholar]
- Immunization of mice with DNA-based Pfs25 elicits potent malaria transmission- blocking antibodies. Infect Immun. 1999;67:1688-93.
- [Google Scholar]
- Sustained high-titer antibody responses induced by conjugating a malarial vaccine candidate to outer-membrane protein complex. Proc Natl Acad Sci USA. 2000;103:18243-8.
- [Google Scholar]
- Development of a Pfs25-EPA malaria transmission blocking vaccine as a chemically conjugated nanoparticle. Vaccine. 2013;31:2954-62.
- [Google Scholar]
- Functional evalution of malaria Pfs25 DNA vaccine by in vivo electroporation in olive baboons. Vaccine. 2013;31:3140-7.
- [Google Scholar]
- A plant-produced Pfs25 VLP malaria vaccine candidate induces persistent transmission blocking antibodies against Plasmodium falciparum in immunized mice. PLoS One. 2013;8(11):e79538.
- [Google Scholar]
- A novel malaria protein, Pfs28, and Pfs25 are genetically linked and synergistic as falciparum malaria transmission-blocking vaccines. Infect Immun. 1997;65:1109-13.
- [Google Scholar]
- Enhanced antibody responses to Plasmodium falciparum Pfs28 induced in mice by conjugation to ExoProtein A of Pseudomonas aeruginosa with an improved procedure. Microbes Infect. 2009;11:408-12.
- [Google Scholar]
- Carboxypeptidases B of Anopheles gambiae as targets for a Plasmodium falciparum transmission-blocking vaccine. Infect Immun. 2007;75:1635-42.
- [Google Scholar]
- Molecular characterization of the carboxypeptidase B1 of Anopheles stephensi and its evaluation as a target for transmission-blocking vaccines. Infect Immun. 2013;81:2206-16.
- [Google Scholar]
- 3’-RACE Amplification of aminopeptidase N gene from Anopheles stephensi applicable in transmission blocking vaccines. Avicenna J Med Biotechnol. 2012;4:131-41.
- [Google Scholar]
- Antibodies to a single, conserved epitope in Anopheles APN1 inhibit universal transmission of Plasmodium falciparum and Plasmodium vivax malaria. Infect Immun. 2014;82:818-29.
- [Google Scholar]
- Expression, immunogenicity, histopathology, and potency of a mosquito-based malaria transmission-blocking recombinant vaccine. Infect Immun. 2012;80:1606-14.
- [Google Scholar]
- Disruption of Plasmodium falciparum development by antibodies against a conserved mosquito midgut antigen. Proc Natl Acad Sci USA. 2007;104:13461-6.
- [Google Scholar]
- PfCCp proteins of Plasmodium falciparum: gametocyte-specific expression and role in complement-mediated inhibition of exflagellation. Int J Parasitol. 2008;38:327-40.
- [Google Scholar]
- A member of the CPW-WPC protein family is expressed in and localized to the surface of developing ookinetes. Malar J. 2013;12:129.
- [Google Scholar]
- Antibodies to plant-produced Plasmodium falciparum sexual stage protein Pfs25 exhibit transmission blocking activity. Hum vaccine. 2011;7(Suppl):191-8.
- [Google Scholar]
- A novel plant-produced Pfs25 fusion subunit vaccine induces long-lasting transmission blocking antibody responses. Hum Vaccin Immunother. 2015;11:124-32.
- [Google Scholar]
- Heat-precipitation allows the efficient purification of a functional plant-derived malaria transmission-blocking vaccine candidate fusion protein. Biotechnol Bioprocess Eng. 2015;112:1297-305.
- [Google Scholar]
- Heterologous expression of the C-terminal antigenic domain of the malaria vaccine candidate Pfs48/45 in the green algae Chlamydomonas reinhardtii. Appl Microbiol Biotechnol. 2013;97:1987-95.
- [Google Scholar]
- Alga-produced cholera toxin-Pfs25 fusion proteins as oral vaccines. Appl Microbiol Biotechnol. 2013;79:3917-25.
- [Google Scholar]
- Algae-produced malaria transmission-blocking vaccine candidate Pfs25 formulated with a human use-compatible potent adjuvant induces high affinity antibodies that block Plasmodium falciparum infection of mosquitoes. Infect Immun. 2015;83:799-808.
- [Google Scholar]
- Plasmodium falciparum: natural and experimental transmission-blocking immunity. Immunol Lett. 1988;19:235-40.
- [Google Scholar]
- Gametocytes of Plasmodium falciparum: phagocytosis by leucocytes in vivo and in vitro. Trans R Soc Trop Med Hyg. 1976;70:344-5.
- [Google Scholar]
- Leukocytes in a Plasmodium falciparum-infected blood meal reduce transmission of malaria to Anopheles mosquitoes. Infect Immun. 1197;65:3834-7.
- [Google Scholar]
- Development and characterization of novel carrier gel core liposomes based transmission blocking malaria vaccine. J Control Release. 2009;140:157-65.
- [Google Scholar]
- Single-dose microparticle delivery of a malaria transmission-blocking vaccine elicits a long-lasting functional antibody response. Curr Mol Med. 2013;13:479-87.
- [Google Scholar]
- Phase 1 trial of malaria transmission blocking vaccine candidates Pfs25 and Pvs25 formulated with montanide ISA 51. PLoS One 2008:e2636.
- [Google Scholar]
- Phase I trial of Pfs25-EPA/Alhydrogel® a transmission blocking vaccine against falciparum malaria in healthy malaria-naïve adults. American Society of Tropical Medicine and Hygiene 61st annual meeting. 2012. p. :441. Abstract#1455
- [Google Scholar]




