Translate this page into:
In vivo oxalate degradation by liposome encapsulated oxalate oxidase in rat model of hyperoxaluria
Reprint requests: Dr C.S. Pundir, Department of Biochemistry, MD University, Rohtak 124 001, India e-mail: pundircs@rediffmail.com
-
Received: ,
This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-Share Alike 3.0 Unported, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This article was originally published by Medknow Publications & Media Pvt Ltd and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Background & objectives:
High level of urinary oxalate substantially increases the risk of hyperoxaluria, a significant risk factor for urolithiasis. The primary goal of this study was to reduce urinary oxalate excretion employing liposome encapsulated oxalate oxidase in animal model.
Methods:
A membrane bound oxalate oxidase was purified from Bougainvillea leaves. The enzyme in its native form was less effective at the physiological pH of the recipient animal. To increase its functional viability, the enzyme was immobilized on to ethylene maleic anhydride (EMA). Rats were injected with liposome encapsulated EMA- oxalate oxidase and the effect was observed on degradation of oxalic acid.
Results:
The enzyme was purified to apparent homogeneity with 60-fold purification and 31 per cent yield. The optimum pH of EMA-derivative enzyme was 6.0 and it showed 70 per cent of its optimal activity at pH 7.0. The EMA-bound enzyme encapsulated into liposome showed greater oxalate degradation in 15 per cent casein vitamin B6 deficient fed rats as compared with 30 per cent casein vitamin B6 deficient fed rats and control rats.
Interpretation & conclusions:
EMA-oxalate oxidase encapsulated liposome caused oxalate degradation in experimental hyperoxaluria indicating that the enzyme could be used as a therapeutic agent in hyperoxaluria leading to urinary stones.
Keywords
Enzyme therapy
hyperoxaluria
liposome
oxalate degradation
oxalate oxidase

Oxalic acid is found in a variety of plants in the form of Ca2+, Mg2+, Na+ and K+ oxalates and, therefore, a component of many constituent in human diet1. Oxalate is not further metabolized in human and is excreted through urine. When combined with calcium, oxalic acid produces an insoluble salt calcium oxalate, which is the most prevalent compound found in kidney stones. High intake of dietary oxalate may limit absorption of calcium and other minerals, which finally leads to hyperoxaluria2. Hyperoxaluria is one of the main risk factors of recurrent urolithiasis and progressive nephrocalcinosis. Oxalate level in an individual is normally checked by a balance between absorption and excretion of dietary oxalates.
In 1955, the discovery of substrate-specific, oxalate degrading enzyme, oxalate oxidase provided a new direction to the research of oxalate stone disease3. Existing methods to treat elevated oxalate level are not always effective. Intensive dialysis, organ transplantation, surgery are not required in patients suffering from primary hyperoxaluria. Current treatment options in patients with primary hyperoxaluria and secondary hyperoxaluria are limited and do not always lead to sufficient reduction in urinary oxalate excretion. Oxalate degrading enzymes have several potential applications, including treatment of hyperoxaluria and other oxalate related disease4. Oxalate oxidase also plays an important role in degradation of oxalic acid in experimental hyperoxaluria5. It has been found in several species viz. bacteria Pseudomonas OX-536, fungus Tilletia controversa7 and in higher plants such as leaves of Bougainvillea spectabilis8, barley seedlings9, Sorghum leaves10, banana fruit peel11, Amaranthus leaves12, wheat seedlings13, maize roots14 and in fruits of strawberry15.
Enzymes in their native forms become inactivated in human body due to proteases and provoke immune reactions16. A most promising development in the field of enzyme therapy has been the use of artificial lipid vesicle for the entrapment of enzymes and for their delivery to cells. These biodegradable liposomes can provide sufficiently large amount of highly purified enzyme to be protected in the circulation for a longer period of time17.
We have earlier purified and characterized a membrane bound oxalate oxidase from Bougainvillea glabra leaves18. The present study was undertaken to examine the possibilities of this enzyme as enzyme supplementation therapy for experimental hyperoxaluria leading to urolithiasis using rat model.
Material & Methods
Oxalic acid, Sephadex G-100, DEAE-Sephacel, 4-aminophenazone, polyvinyl polypyrrolidone (PVPP), taurodeoxycholate sodium salt were purchased from Sigma Aldrich, St. Louis. USA. Horse radish peroxidase (HRP, RZ 3.0), chloroform, cholesterol were purchased from SISCO Research laboratory Pvt. Ltd, Mumbai. L-α phosphatidyl choline was purchased from Hi-media, Mumbai, India. 14C-Oxalic acid (50 μCi) was from American Radiolabelled Chemicals (ARC, St. Louis, USA). All other chemicals were of analytical reagent (AR) grade, ethylene maleic anhydride (EMA) was purchased from Vertillus, USA.
The study was conducted in the department of Biochemistry, M.D. University, Rohtak, India.
Preparation of B. glabra oxalate oxidase: The experiment was carried out at 4°C according to method19 with a few modifications. Mature leaves of B. glabra (200 g) were collected from M.D. University campus, Rohtak, and homogenized in an extraction medium [0.1 M potassium phosphate buffer, pH 7.0 containing 0.5 M sucrose, in a ratio of 1:5 (w/v)] and 1 per cent polyvinyl polypyrrolidone (PVPP). The leaf homogenate was filtered through three folds of cheese cloth and filtrate was centrifuged at 15,000×g for 30 min at 4°C and pellet was collected and suspended in a minimum quantity of extraction medium and treated as crude enzyme.
Solubilization of membrane bound oxalate oxidase from leaves of B. glabra: The solubilization of membrane bound Bougainvillea leaf oxalate oxidase was achieved20. Solid taurodeoxycholic acid was added (at concentration 5-20 mg/ml), kept under constant stirring at 4°C for different time periods. The suspension containing solubilized enzyme was centrifuged at 15,000×g for 30 min. Both the supernatant and pellet were collected and assayed for activity using standard assay conditions and protein by Lowry method21. The supernatant was treated as crude enzyme and subjected to purification using the following steps:
Ammonium sulphate precipitation: The crude extract was used for an ammonium sulphate at 0-80 per cent saturation. The precipitate was collected by centrifugation at 12,000×g for 30 min and suspended in minimum quantity of extraction medium.
Gel filtration on Sephadex G-100 column: The pellets obtained from ammonium sulphate precipitation were applied to Sephadex G-100 (21×2 cm) column previously equilibrated with 0.02 M potassium phosphate buffer (pH 6.8). The activity of enzyme and protein content in fraction (3 ml each) were measured. The fractions containing high specific activity were pooled.
Ion-exchange chromatography on DEAE-Sephacel: The pooled fraction of Sephadex G-100 was loaded on DEAE-Sephacel column (20×0.5 cm) along its side wall. The enzyme was eluted in linear gradient of KCl (0.1 M-0.6 M) using 100 ml 0.02 M potassium phosphate buffer, pH 6.8 containing 0.1 M KCl in a mixer and 100 ml of the same buffer in 0.6 M KCl in reservoir. The buffer in mixing aspirator was stirred with the help of a magnetic stirrer. The flow rate was maintained at 0.5 ml per min. The fractions (3 ml each) were collected and monitored for enzyme activity and protein content. The fractions with high specific activity were pooled and treated as purified enzyme.
Assay of oxalate oxidase: Assay of oxalate oxidase was carried out as described elsewhere8 with certain modifications. The reaction mixture containing 1.8 ml of 0.05 M sodium succinate buffer pH 5.5, 0.1 ml of enzyme was pre-incubated at 37°C for 2 min. The reaction was started by adding 0.1 ml of oxalate (0.01 M). After incubation at 40°C in dark for 4 min, 1.0 ml colour reagent (50 mg 4-aminophenazone, 100 mg solid-phenol and 1mg horse radish peroxidase per 100 ml of 0.4 M sodium-succinate buffer, pH 7.0, stored in amber coloured bottle at 4°C and prepared fresh every week) was added and kept at room temperature (25±5°C) in dark to develop colour. In blank, the enzyme was replaced by reaction buffer. Absorbance of the reaction mixture was read at 520 nm in a spectrophotometer (Spectronic -20, USA). H2O2 generated in the assay was calculated from standard curve between H2O2 conc. vs. A520.
One unit of enzyme is defined as the amount of enzyme required to catalyze the generation of 1 μmoles of H2O2 from oxalate per min under standard assay conditions.
Determination of protein concentration: Protein concentration in various enzyme preparations was determined by Lowry method21 using bovine serum albumin (BSA) as standard protein.
The purity of enzyme was tested by PAGE using coomasie blue as protein stain.
Formation of ethylene maleic anhydride (EMA) derivative oxalate oxidase: The cold freshly prepared solution of enzyme in 0.02 M potassium phosphate buffer, pH 7.0, was slowly added to the magnetically suspended EMA derivative (100 mg in 5 ml of same buffer). The reaction mixture was left stirring overnight at 4°C. The water insoluble EMA-derivative was separated by centrifugation at 12,000×g for 30 min and washed with distilled water to get EMA-derivative oxalate oxidase.
Liposome preparation and encapsulation of EMA- oxalate oxidase: Liposomes were prepared according to the method by Gregoriadis et al22 with modification. Briefly, egg phosphatidylcholine (10 mg) and cholesterol (1 mg) were dissolved in 1 ml of chloroform, which was subsequently eliminated by rotatory evaporation under reduced pressure. The process was allowed to continue till liquid was evaporated completely and a dry lipid film was formed on the side walls of the flask. The flask was rotated under vacuum flushed with nitrogen for another 15 min to remove last traces of solvent. The thin film formed on the walls of the flask was dispersed with 0.02 M sodium phosphate buffer, pH 7.0 containing 400 unit of EMA derivative oxalate oxidase, which was kept for 1-2 h for complete swelling. The free non-entrapped oxalate oxidase enzyme was separated from encapsulated oxalate oxidase liposome by centrifugation at 15,000×g for 1 h at 4° C in a refrigerated centrifuge. The pellets formed were dissolved in 0.02 M sodium phosphate buffer and kept at 4°C until use. The photomicrograph of liposomes were taken with a projection microscope fitted with CCD camera (Micron optic microscope, IS: 4381, ISI, Model-TMC-II, USA) under 100x magnification.
Determination of encapsulation efficiency: The percentage of enzyme encapsulated was determined by centrifugation as described by Panwar et al23 with modifications. Aliquots of liposomal dispersion were subjected to centrifugation at 15,000×g for 1 h. The clear supernatant was removed carefully to separate non-entrapped oxalate oxidase and absorbance was recorded at 520 nm. The sediment containing entrapped oxalate oxidase was diluted to 100 ml with potassium phosphate buffer, pH 7.4 and its absorbance was recorded at 520 nm.
Animal experiment: This study was approved by the ethical committee of MDU, Rohtak. Twelve male albino rats (weighing approx 100 g) were used and randomized to three groups kept in the animal house of M.D. University, Rohtak. One group was fed a standard diet with vitamin B6 (control), while the other rats were fed with 15 per cent casein vitamin B6 deficient diet (test group B) and 30 per cent casein vitamin B6 deficient diet (test group C) for 20 wk24–26.
Determination of oxalate content in control and test rats: The first morning urine oxalate content was determined by enzymatic method10 with some modifications. Urine of control and test rats was collected in a vial containing 1.5 ml concentrated HCl. To the acidified urine, 1.0 ml of 0.2 mol/l NaOH was added to increase pH to 6.0 followed by addition of 0.1 ml NaNO2 (35 mg/10 ml). The pretreated urine was used for the determination of oxalate content. The oxalate content was determined in the similar manner as described for assay of oxalate oxidase except that oxalate was replaced by pretreated urine samples and optimal assay conditions were maintained. The concentration of oxalate in urine was extrapolated from standard curve drawn from oxalate concentration ranging from 0.001 mM to 0.07 mM and A520.
Study of effect of liposome encapsulated EMA-bound oxalate oxidase on degradation of oxalic acid: To study the effect of liposome encapsulated EMA-oxalate oxidase in degradation of oxalate, test rats groups B and C (15 and 30% casein diet) were intraperitoneally injected with liposome encapsulated oxalate oxidase. Adult female rats were given diet of evaporated milk and water prior to each experiment27. To the individual rats 0.6 ml of 3.6 mg labelled 14C Oxalic acid in sodium phosphate buffer was administered intraperitoneally. Control rats received the same treatment except intraperitoneal injection of liposome encapsulated oxalate oxidase. The treated rats were housed in cages for urine collection. The amount of unmetabolized oxalate was extrapolated from the standard curve between amount of radiolabelled [14C] oxalate and cpm.
Statistical analysis: The values were the mean of 3 replications.
Results & Discussion
In the present study, the membrane bound oxalate oxidase was purified from mature leaves of B. glabra by 60-fold with a specific activity 20 unit/mg protein confirming our earlier observations15.
Effect of pH on EMA bound enzyme: The pH profile of free oxalate oxidase and EMA-oxalate oxidase are shown in Fig. 1. The native oxalate oxidase exhibited optimal activity at pH 5.5, the enzyme showed only 55 per cent activity at pH 7.0. However, EMA bound enzyme showed a shift in its optimum pH from pH 5.5 to pH 6.0. At pH 7.0, EMA-oxalate oxidase displayed 72 per cent activity.
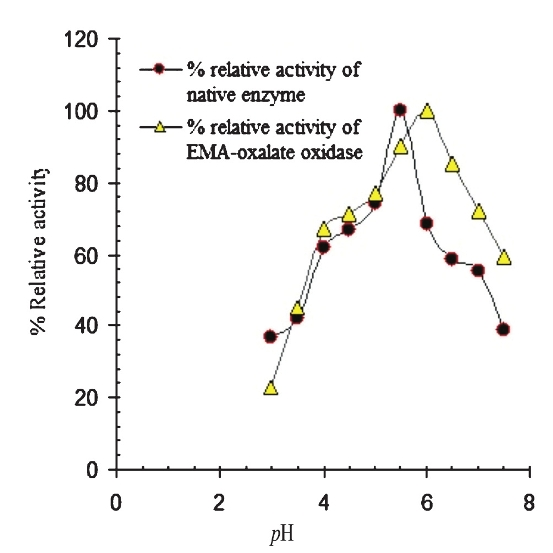
-
pH profile of native and EMA-oxalate oxidase purified from Bougainvillea glabra leaf.
Liposome preparation and encapsulation of EMA-oxalate oxidase: Enzyme encapsulated liposome were prepared by combination of phosphatidylcholine: cholesterol in the ratio of 10:1 dissolved in chloroform solution. Photomicrograph structure clearly shows the formation of liposome. Most of the liposome formed were spherical, heterogeneous and unilamellar in arrangement (Fig. 2 a and b). The encapsulation efficiency was found to be 74 per cent at above ratio of phosphatidyl choline and cholesterol.
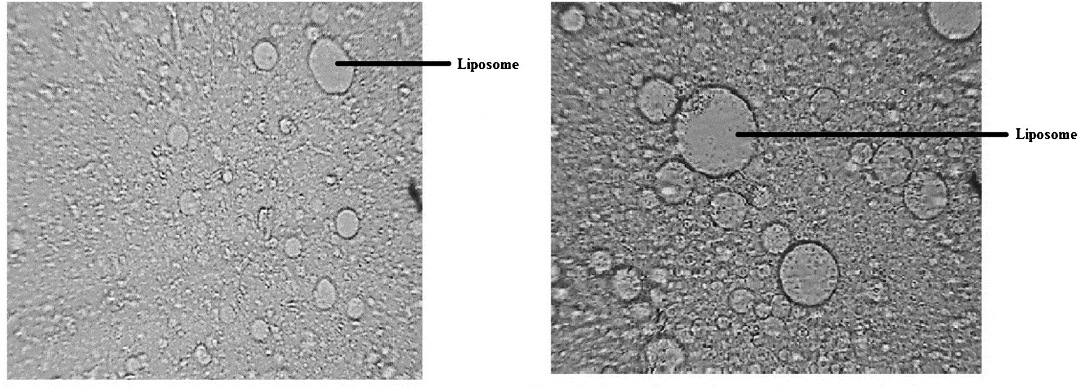
- Photomicrograph of liposome formed by mixing phosphtidyl choline and cholesterol. (a) 100 × (b) 200 ×.
Determination of oxalate in urine of control and test rats: In vitamin B6 deficient rats, there was a marked increase in urinary oxalate excretion compared to control (P<0.001). Oxaluria was more pronounced in 30 per cent casein test group rats as compared with control. The urinary oxalate content in control rats ranged from 0.48-0.85 mg/24 h in 20 wk fed with normal diet, whereas in 15 per cent hyperoxaluric rats, the oxalate content ranged from 1.8-3 mg/24 h in 20 wk, when they were fed with 15 per cent casein diet. The oxalate content in 30 per cent casein fed rats was increased 4-fold compared with control, as it ranged from 2-4 mg/24 h (Fig. 3). These results showed that the urinary oxalate excretion was increased significantly in both 30 and 15 per cent casein (vit. B6 deficient) fed test rats, compared to control (P<0.001).
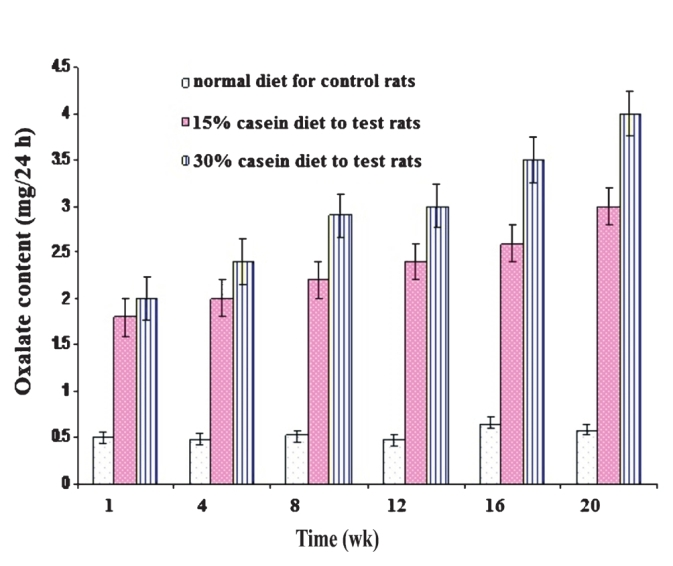
- Effect of vitamin B6 deficiency on urinary oxalate excretion in albino adult male rats. Oxalate excretion significantly higher in both 30-15 per cent casein (vitamin B6 deficient) fed rats compare to control (P<0.001). Values are mean ±SD of n=3 experiments.
Enzymatic degradation of oxalate in hyperoxaluric rats: The intraperitoneal injection of liposome encapsulated EMA-oxalate oxidase resulted into higher rate of metabolic degradation of administered [14C] oxalic acid. Reduction in the functional performance of liposome encapsulated oxalate oxidase was observed in 30 per cent casein fed rats as compared with 15 per cent casein group rats. This may be due to loss of activity of oxalate oxidase to act on higher concentration of oxalic acid. Thus compared with control, oxalate oxidase treated animals excreted lesser amount of unmetabolized [14C] oxalic acid. Control rats were able to excrete 88-92 per cent of injected oxalate, whereas oxalate oxidase excreted merely 44 per cent of injected oxalate in 15 per cent casein fed B6 deficient rats and 52.7 per cent in 30 per cent deficient casein fed rats (Fig. 4).
![Urinary oxalate excretion in 24 h urine in control and test rats by daily injection of [14C] oxalic acid. Inset shows the standard curve of radiolabelled [14C] oxalic acid vs cpm (P<0.001).](/content/175/2013/137/1/img/IJMR-137-136-g004.png)
- Urinary oxalate excretion in 24 h urine in control and test rats by daily injection of [14C] oxalic acid. Inset shows the standard curve of radiolabelled [14C] oxalic acid vs cpm (P<0.001).
In conclusion, our results showed the in vivo degradation of oxalate by exogenously injected liposome encapsulated EMA bound oxalate oxidase under experimental hyperoxaluria. These results indicate that the encapsulated Bougainvillea leaf oxalate oxidase was able to degrade oxalate in rat model with experimental hyperoxaluria, which lead to urinary stone. However, further studies need to be done to confirm these findings.
Acknowledgment
Authors thank Drs Balraj Singh and P. Mookherjee, NRL, IARI, New Delhi, for providing liquid scintillation counting facility for labelled 14C-oxalic acid and Dr Arun Nanda, Department of Pharmaceutical Science, M.D. University, Rohtak, for providing projection microscope used in liposome study.
References
- Oxalic acid in biology and medicine. London: Academic Press; 1977.
- Hyperoxaluria in L-glyceric aciduria: possible nonenzymic mechanism. Biochem Med. 1983;29:114-21.
- [Google Scholar]
- Isolation, purification, immobilization of oxalate oxidase and its clinical applications. Hindustan Antibiot Bull. 1993;35:173-82.
- [Google Scholar]
- Degradation of oxalate in rats implanted with immobilized oxalate oxidase. FEBS Lett. 1986;195:101-5.
- [Google Scholar]
- Purification and characterization of oxalate oxidase from Pseudomonas sp. OX-53. Agric Biol Chem. 1988;52:743-8.
- [Google Scholar]
- Oxalate oxidation by an obligately parasitic fungus Tilletia contraversa. Arch Biochem Biophys. 1961;95:66-9.
- [Google Scholar]
- An oxalic acid oxidase in the leaves of Bougainvillea spectabilis. Biochem J. 1962;85:33-8.
- [Google Scholar]
- Purification and properties of oxalic acid oxidase. Arch Biochem Biophys. 1966;116:516-23.
- [Google Scholar]
- Occurrence of an oxalate oxidase in Sorghum leaves. Phytochemistry. 1984;23:1871-4.
- [Google Scholar]
- Oxalate oxidase from banana peel for determination of urinary oxalate. Clin Chem. 1985;31:649.
- [Google Scholar]
- Purification and properties of a membrane bound oxalate oxidase from Amaranthus leaves. Plant Sci. 1999;142:21-8.
- [Google Scholar]
- Germin, a protein marker of early plant development is an oxalate oxidase. J Biol Chem. 1993;268:12239-42.
- [Google Scholar]
- Characterization of cell wall oxalate oxidase from maize roots. Plant Sci. 2000;157:257-63.
- [Google Scholar]
- Strawberry fruit oxalate oxidase: detection, purification characterization and physiological role. J Plant Biochem Biotech. 2010;19:247-50.
- [Google Scholar]
- Desnick RJ, ed. Enzyme therapy in genetic diseases. Vol 2. New York: Alan R. Liss; 1980.
- Physiological pharmaceutics: barriers to drug absorption (2nd ed). London: Taylor and Francis; 2001.
- Purification and properties of membrane bound oxalate oxidase from leaves of Bougainvillea glabra. Int J Biotechnol Biochem. 2011;7:515-28.
- [Google Scholar]
- Comparative study of oxalate oxidase in three genotypes of Sorghum vulgare. Experientia. 1991;47:599-602.
- [Google Scholar]
- Quantification of urinary oxalate with oxalate oxidase from beet stems. Clin Chem. 1983;29:1815-9.
- [Google Scholar]
- Comparative effect and fate of non-entrapped and liposome-entrapped neuraminidase injected into rats. Biochem I. 1974;140:323-30.
- [Google Scholar]
- Preparation, characterization, and in vitro release study of albendazole- encapsulated nanosize liposome. Int J Nanomedicine. 2010;5:101-8.
- [Google Scholar]
- Endogenous oxalate synthesis and glycine, serine, deoxypyridoxine interrelationships in vitamin B6-deficient rats. J Biol Chem. 1959;234:2391-3.
- [Google Scholar]
- Effect of high protein diet on stone-forming propensity and bone loss in rats. Kidney Int. 2003;64:2142-9.
- [Google Scholar]
- Comparative structural modeling and docking studies of oxalate oxidase: possible implication in enzyme supplementation therapy for urolithiasis. Int J Biol Macromol. 2011;48:466-73.
- [Google Scholar]
- The metabolism of ascorbic acid -1-14C and oxalic acid -C14 in the rat. J Biol Chem. 1955;216:539-48.
- [Google Scholar]




