
Translate this page into:
Lymphopenia-induced proliferation of CD4 T-cells is associated with CD4 T-lymphocyte exhaustion in treated HIV-infected patients
For correspondence: Dr. Evgeniya V. Saidakova, Institute of Ecology & Genetics of Microorganisms, UB RAS, 614081, Perm, 13 Goleva Str., Russia e-mail: radimira@list.ru
-
Received: ,
This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
This article was originally published by Medknow Publications & Media Pvt Ltd and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Background & objectives:
Under the lymphopenic condition, T-cells divide to maintain their peripheral pool size. Profound chronic lymphopenia in some treated HIV-infected patients, characterized by poor T-cell recovery, might result in intensive homeostatic proliferation and can cause T-cell exhaustion and/or senescence. The present study was undertaken to evaluate the homeostatic proliferation of CD4+ T-cells in treated HIV-infected individuals, and to determine the amount of phenotypically exhausted and senescent CD4 T-lymphocytes.
Methods:
Thirty seven treated HIV-infected patients with suppressed HIV viral load (<50 copies/ml) were studied. Patients were divided into two groups: immunological non-responders (INRs) with CD4+ T-cells <350/μl (n=16) and immunological responders (IRs) with CD4+ T-cells >350/μl (n=21). T-cell subsets [naïve, central memory (CM), and effector memory (EM)] and proportions of cycling (Ki-67+), senescent (CD57+) and exhausted (PD-1+) T-lymphocytes were assessed using flow cytometry.
Results:
CD4+ T-cell cycling rate was higher in INRs than in IRs due to more extensive proliferation of CM, 4.7 vs 2.7 per cent (P <0.01) and EM, 4.8 vs 3.2 per cent (P <0.05). The percentages of CD4+ Ki-67+ CM and EM T-lymphocytes were inversely related to the CD4+ T-cell counts in the appropriate subset, r=–0.584 (P <0.001) and r=–0.556, (P <0.001), respectively. Exhaustion [24.2 vs 16.7% (P <0.01)], but not senescence [7.1 vs 10.8% (P>0.05)] was more pronounced in the INR group than in the IR group. The frequency of CD4+ Ki-67+ CM T-cells was related to the proportion of CD4+ PD-1+ cells of the same subset, r=0.789 (P <0.001). The numbers of CD4+ Ki-67+ PD-1+ CM and EM T-cells were substantially higher in INRs than in IRs.
Interpretation & conclusions:
The present data indicated that intensive homeostatic proliferation contributed to the T-cell exhaustion in HIV-infection.
Keywords
CD57
exhaustion
HIV infection
lymphopenia
PD-1
proliferation
T cell

Homeostatic T-cell proliferation occurs in lymphopenic conditions12. This type of division takes place at a polyclonal level and does not require any foreign antigen3. It applies to both naive and memory T-cells and is driven by cytokines, such as interleukin (IL)-7 and IL-15, and low-affinity interactions with self-peptides loaded on the major histocompatibility complex (MHC) molecules45. In HIV-infection, T-cell depletion is associated with homeostatic T-cell response. In particular, CD4+ T-cell count has been shown to be inversely correlated with the number of CD4+ cycling T-cells67. Lymphopenic immunological non-responders (INRs), HIV-infected patients on effective highly active antiretroviral therapy (HAART) with non-detectable viral load (<50 copies/ml) who fail to restore their CD4+ T-cell count were shown to have higher rates of homeostatic proliferation than those responding to treatment with successful reconstitution of their CD4+ T-cell pool8. While it is established that intensive proliferation can be a reason for the formation of the T cell pool with compromised functional activity9, the role of homeostatic division in the development of functionally impaired T-cells in HIV-infected INR patients remains to be determined.
Exhaustion and senescence are the two states that are characterized by the impairment of T-cell functions1011. Immune exhaustion is a progressive decrease in the reactivity of T-cells during long time antigen persistence12. One of the molecules specific for functional T-cell exhaustion is programmed death-1 (PD-1) protein which conveys inhibitory signals to T-lymphocytes after the interaction with the appropriate ligands1314. PD-1 is significantly upregulated on HIV-specific CD4+ and CD8+ T-cells during the infection15. Its expression correlates with impaired T-lymphocyte functions as well as with indices of HIV-infection progression: positively with HIV viral load and inversely with CD4 T-cell count15. Furthermore, PD-1/PD-L pathway blockade was shown to reinvigorate the exhausted T-cells in vitro suggesting that exhaustion might directly inhibit immune functions101617. High percentage of PD-1-positive T-cells was identified as a unique marker in the HIV-infected INR patients16.
The senescent state of T-cells develops as a result of many rounds of proliferation9. The molecule specific to senescent T cells is CD57. Its expression correlates with impairment in IL-2 producing activity and predisposition to apoptosis11. HIV-infection is characterized by the large proportion of senescent T cells18, but little is known about the influence of homeostatic proliferation on the formation of the senescent T cell pool in INR patients and the role of senescence in immunological non-response to HAART.
To characterize the effect of lymphopenia-induced cell division on major peripheral CD4 T-cell maturational subsets of HIV-infected individuals, the homeostatic proliferation in treated HIV-infected patients with low and close to normal T-cell counts was evaluated. Furthermore, the amounts of phenotypically exhausted and senescent CD4 T-lymphocytes were also estimated.
Material & Methods
This study was approved by the Institutional Review Board of the Perm Regional Center for Protection against AIDS and Infectious Diseases, Perm, Russia (IRB00008964). Each participant provided a written informed consent. The work was carried out at the Laboratory of Ecological Immunology, Institute of Ecology and Genetics of Microorganisms, Ural Branch, Russian Academy of Sciences (UB RAS) and Perm State University, Perm, Russia between August 2012 and April 2013.
Eighty HIV-infected patients [38 INRs & 42 immunological responders (IRs)] were chosen from the electronic database of the Medical Assistance Department in the Perm Regional Center for Protection against AIDS and Infectious Diseases. The inclusion criteria were as follows: age over 18 yr, established chronic HIV-infection (confirmed by the ELISA, Western blot & PCR assays), high adherence to the HAART regimen (two nucleoside reverse transcriptase inhibitors with a ritonavir-boosted protease inhibitor or a non-nucleoside reverse transcriptase inhibitor) for more than two years, and HIV viral load suppressed to the undetectable level (<50 copies/ml). Exclusion criteria were as follows: age under 18 yr, pregnancy, active alcohol consumption, active drug usage, diabetes and cancer. To avoid the influence of hepatitis C virus (HCV) infection that is widely spread among the HIV-infected individuals in Russia, 43 HIV/HCV-co-infected patients were excluded from the study. The absence of HCV-co-infection was confirmed by a PCR-based assay (’Quantitative RT-Gepatogen C’ kit; DNA-Technology, Moscow, Russia).
Thirty seven patients who matched all the criteria were divided into two groups (Table) according to the efficacy of their CD4+ T-cell response to HAART. INRs were those who failed to restore their CD4+ T-cells, with <350 CD4+ T-cells/μl of blood, after more than two years of virologically effective HAART (n=16). IRs were those who succeeded in restoring their CD4+ T-cells, more than 350 CD4+ T-cells/μl of blood, after more than two years of virologically effective HAART (n=21).
Patients had no difference in HIV viral load before initiation of HAART, (253285 vs 73800 copies/ml). It was found that the CD4+ T-cell counts before the start of HAART were significantly lower in INR patients than in IR patients [95 vs 160 cells/μl (P <0.01)], confirming that low initial pre-HAART CD4+ T-cell count was a predictor of insufficient immunologic response to the therapy19.
Peripheral blood mononuclear cells (PBMCs): PBMCs were isolated from 30 ml venous blood by Diacoll (ρ=1.077; Dia-M, Moscow, Russia) density sedimentation. The obtained cells were cryopreserved in medium containing 90 per cent of foetal calf serum (Gibco, NY, USA) and 10 per cent of dimethyl sulphoxide (Darmstadt, Germany) and stored at –196°C for six months.
Flow cytometry: Cells were thawed and stained with a set of monoclonal antibodies: anti-CD3-PerCP, anti-CD4-AF700, anti-CD27-APC-Cy7, anti-CD45RA-APC, anti-CCR7-PE-Cy7, anti-Ki-67-PE, anti-HLA-DR-FITC, anti-CD38-PE (Becton Dickinson, USA), anti-PD-1-BV421 (Biolegend, USA), as well as with the LIVE/DEAD® Fixable Yellow Dead Cell Stain Kit (Invitrogen, USA). The analysis was performed using BD LSR II Flow Cytometer (BD Biosciences, USA). Naïve T-cells were identified as CD27+ CD45RA+ CCR7+, central memory (CM) T-cells as CD27+ CD45RA– CCR7+ and effector memory (EM) T-cells as CD27– CD45RA– CCR7–20. Ki-67+ T-cells were considered to be proliferating, PD-1+ T-cells were considered to be exhausted and CD57+ T-cells were considered to be senescent. HLA-DR+ CD38+ T-cells were counted as activated. At least 100,000 events in the lymphocyte gate were collected for each sample analysis.
Enzyme Linked Immunosorbent Assay (ELISA): The plasmas were thawed and diluted as needed to assure that results were within the linear range of the assay. The concentrations of IL-1β, IL-6, IL-7 and IL-15 were determined using ELISA kits purchased from R&D Systems (Minneapolis, MN, USA) according to the manufacturer's instructions.
Statistical analysis: All statistical analyses were performed using Stata 11.1 SE software (StataCorp, Texas, USA). No adjustments were used. The groups were compared using the Mann–Whitney U-test. First calculations were accomplished with groups comprised of 38 INR patients and 42 IR patients with power equal to 0.9. After the exclusion of HIV/HCV co-infected patients, the statistical power dropped, but all the results held the same consistent pattern. Data were reported as medians and quartile ranges. Correlation analyses were performed using the Spearman's rank correlation method.
Results
The patients’ ages were comparable between the two groups (Table). The median ages of INRs and IRs were 35 and 34 yr, respectively. Women were overrepresented in both INR and IR groups (75 & 81%, respectively), and the difference in the gender ratio between the groups was not significant. Sexual HIV transmission route prevailed in both groups. The known duration of HIV-infection and HAART were similar in the two groups. The CD4+ T-cell counts remained lower in the INR group than in the IR group during the time of the study, 280 vs 540 cells/μl (P <0.001), respectively.
T-cell subsets: A small number of circulating CD4+ T-lymphocytes in INRs blood was due to a cell deficiency in each cell subset. Naïve T-lymphocyte absolute counts were much lower in the INR group when compared with the IR group [105 vs 235 cells/μl (P <0.001), respectively]. Similar situation was observed in the CD4+ CM [90 vs 141 cells/μl (P <0.001)] and the CD4+ EM [42 vs 86 cells/μl (P=0.001)] compartments in the INR and IR groups, respectively. The proportions of naïve cells were similar in INR and IR groups [35.1 vs 36.3% (P>0.05)]. There were more CM cells in the INR compared to the IR group [35.9 vs 28.8% (P <0.05)]. However, the proportions of CD4 EM cells were the same for INR and IR patients [17.4 vs 13.3%].
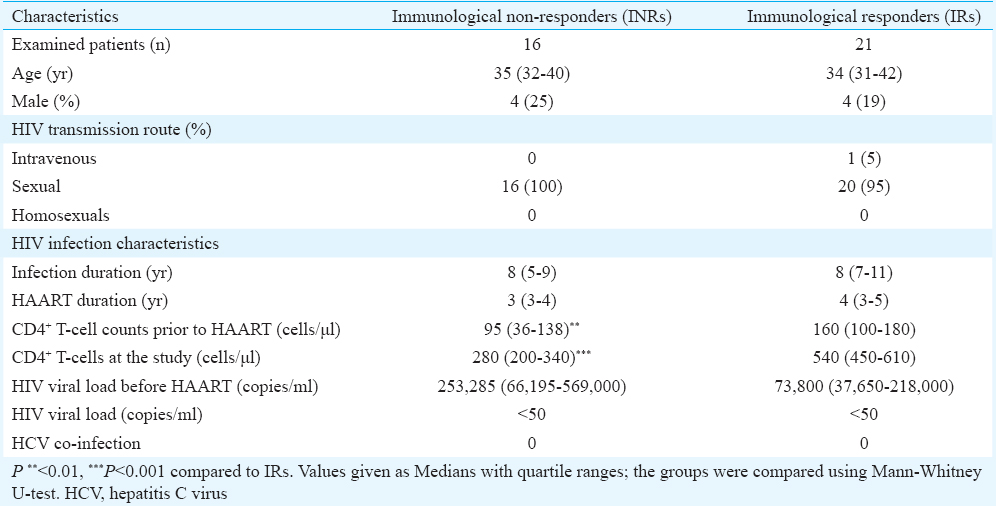
T-cell cycling: During the chronic HIV-infection, different CD4+ T-cell subsets proliferate with a different rate (Fig. 1A). The least vigorous proliferation was observed in the pool of naïve CD4+ T-cells. At the same time, the percentage of Ki-67+ CM CD4+ T-cells was 7.5 times higher than in the pool of naïve CD4+ T-lymphocytes, and the frequency of Ki-67+ EM CD4+ T-cells was more than 10 times higher than the percentage of Ki-67+ naïve CD4+ T-lymphocytes.
- Ki-67 expression on different CD4+ T cell subsets of HIV infected patients. (A) Percentages of Ki-67+ naïve, central memory and effector memory CD4+ T cells in the combined group (INR+IR; n=37) of HIV infected patients. Frequencies of proliferating (B) naïve, (C) central memory, and (D) effector memory T cells in INR (n=16) and IR (n=21) groups. Medians and quartile ranges are shown. INR, immunological non-responders; IR, immunological responders. Hollow circles designate outliers; *P<0.05 compared to IRs.
INRs had a higher percentage of proliferating CD4+ T-cells than IRs, [9.3 vs 6.9% (P <0.01)] which was explained by the more extensive cycling of both CD4+ CM and EM T-cell subsets (Fig. 1C & D), as Ki-67 expression did not differ between the naïve CD4+ T-cell compartments of the INR and IR groups (Fig. 1B).
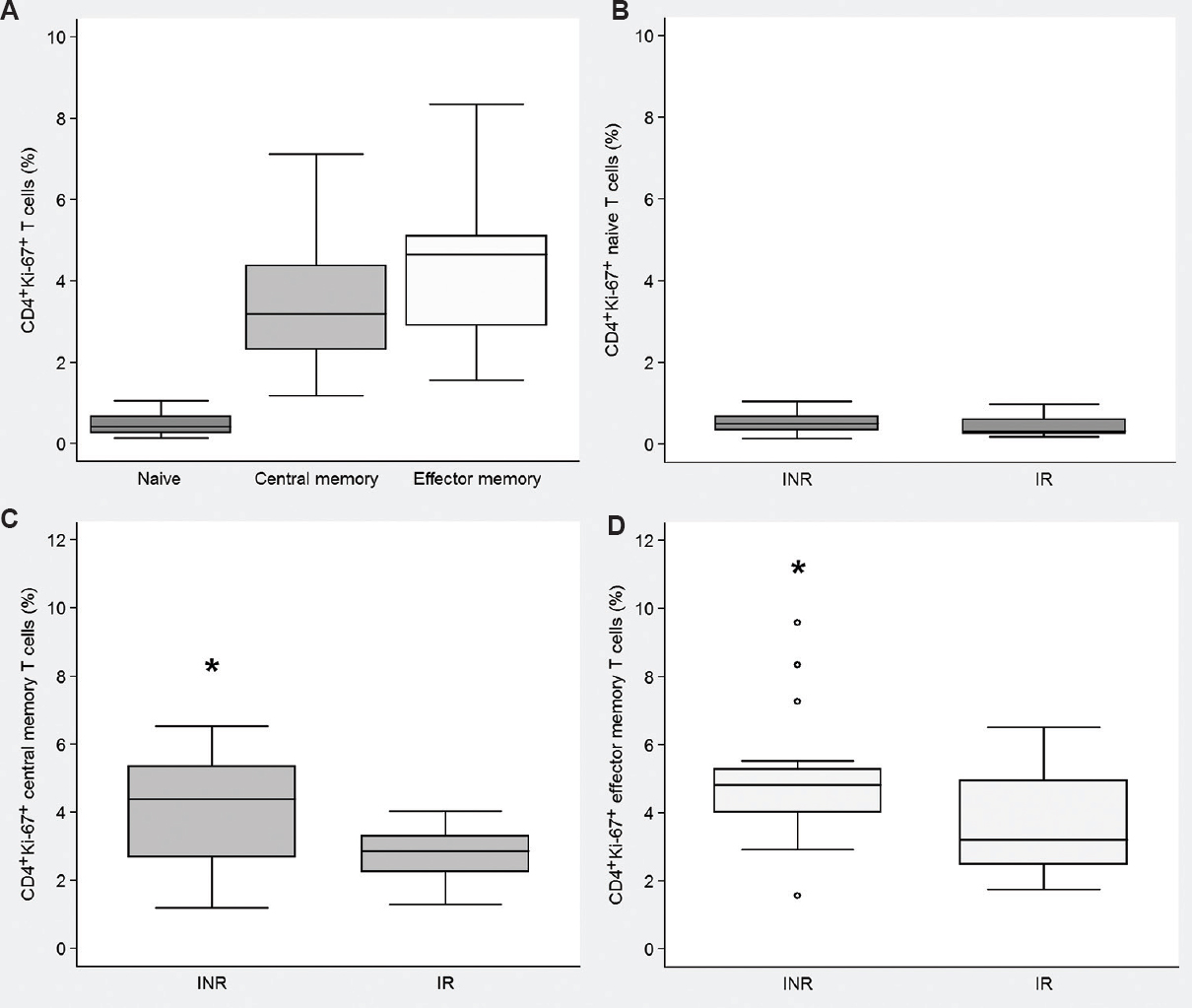
In HIV-infected individuals, the percentage of Ki-67+ CD4+ CM T-cells was inversely correlated with the CD4+ CM T-cell counts, r=−0.584, P <0.001 (Fig. 2A) suggesting the homeostatic processes being the reason for cell division. A similar correlation was observed in the EM CD4+ T-cell compartment, r=−0.556, P <0.001 (Fig. 2B). While the proportions of activated CD4+ HLA-DR+ CD38+ T-cells[12.0 vs 7.0%], levels of IL-1β [0.19 vs 0.14 pg/ml] and IL-6 [0.89 vs 0.94 pg/ml] in plasma were similar in INR and IR groups, respectively, CD4+ T-cell cycling had no correlations with CD4+ T-cell activation and plasma levels of proinflammatory cytokines confirming bystander cell activation and inflammation were not the major reasons for the proliferation observed.
- Correlation between the absolute CD4+ T cell counts and the frequencies of CD4+Ki-67+ T lymphocytes. (A) Central memory subset (n=37). (B) Effector memory subset (n=36).
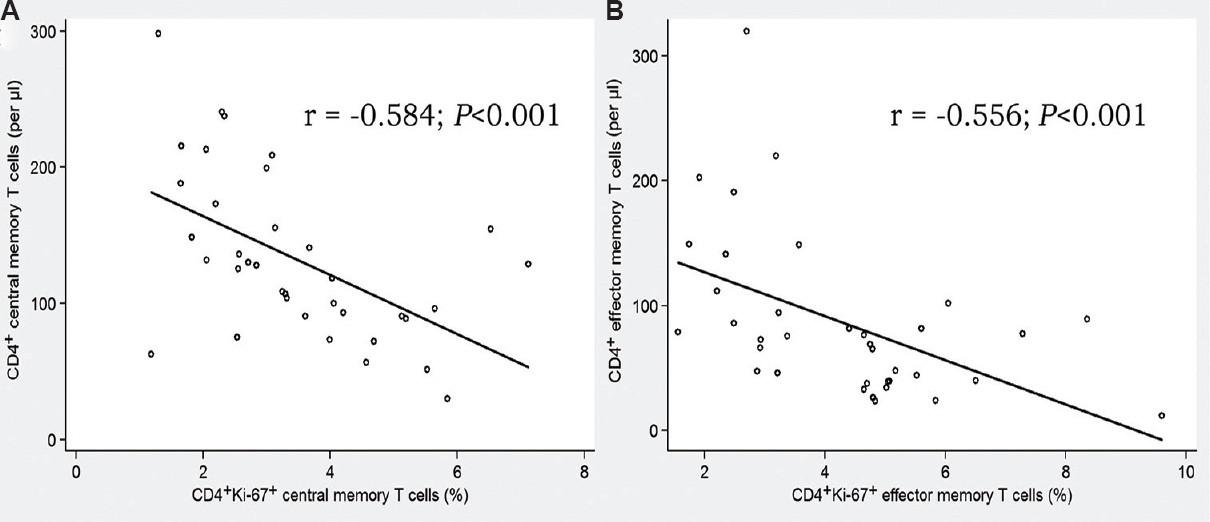
Immune exhaustion and senescence: CD4+ T-cell exhaustion (PD-1 molecule expression) was more pronounced in the INR group than in the IR group, 24.2 vs 16.7 per cent (P <0.01), respectively (Fig. 3A). However, there was no difference in the percentage of CD4+ T-cells expressing CD57 molecule between the INR and IR groups, 7.1 vs 10.8 per cent, respectively (Fig. 3B), supporting the notion that PD-1 and CD57 upregulation in HIV-infection are driven by independent mechanisms.
- Frequencies of exhausted and senescent CD4+ T cells in INR and IR groups. (A) PD-1+ “exhausted” T cells. (B) CD57+ “senescent” T cells. Medians and quartile ranges are shown. INR, immunological non-responders (n=16); IR, immunological responders (n=21). Hollow circles designate outliers. **P<0.01 compared to IR.
It is of importance that during HIV-infection the frequency of proliferating CD4+ CM T-cells was positively correlated with the percentage of the exhausted CD4+ PD-1+ CM T-cells, r=0.789 (P <0.001; Fig. 4). However, such correlation was not observed in the CD4+ EM compartment, r=0.074 (P<0.05). It should be noted that the amount of CD4+ PD-1+ T-cells in HIV-infected people was not dependant on the patients’ age, sex, HIV-infection duration, CD4 T-cell counts before the therapy or HAART duration (data not shown).
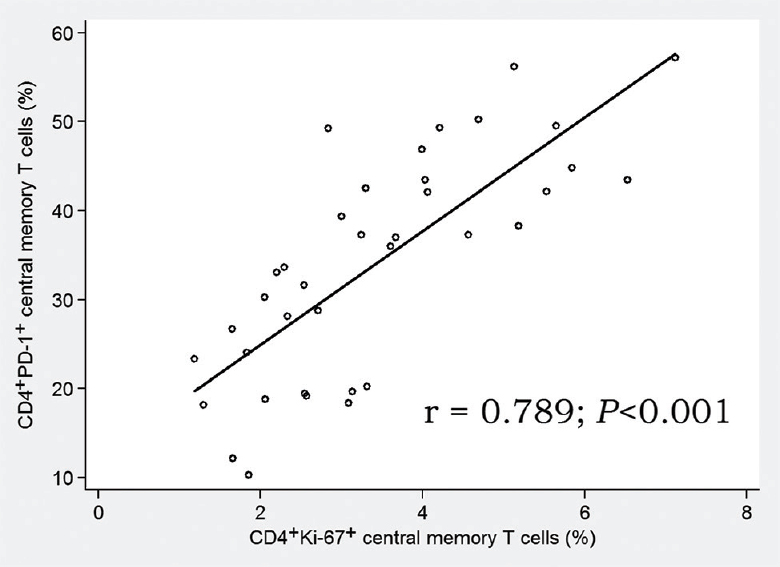
- Correlation between proliferating and exhausted central memory CD4+ T cells. Combined group of HIV infected patients (n=37).
The INRs and the IRs differed in the simultaneous expression of Ki-67 and PD-1 molecules on CD4+ T-cells. In INRs, percentage of CM PD-1+ Ki-67+ CD4+ T-cells was higher than in the IR group, 3.9 vs 2 per cent (P=0.02; Fig. 5A). Similar situation was observed in the EM compartment, 4.3 vs 2.3 per cent (P <0.01; Fig. 5B).
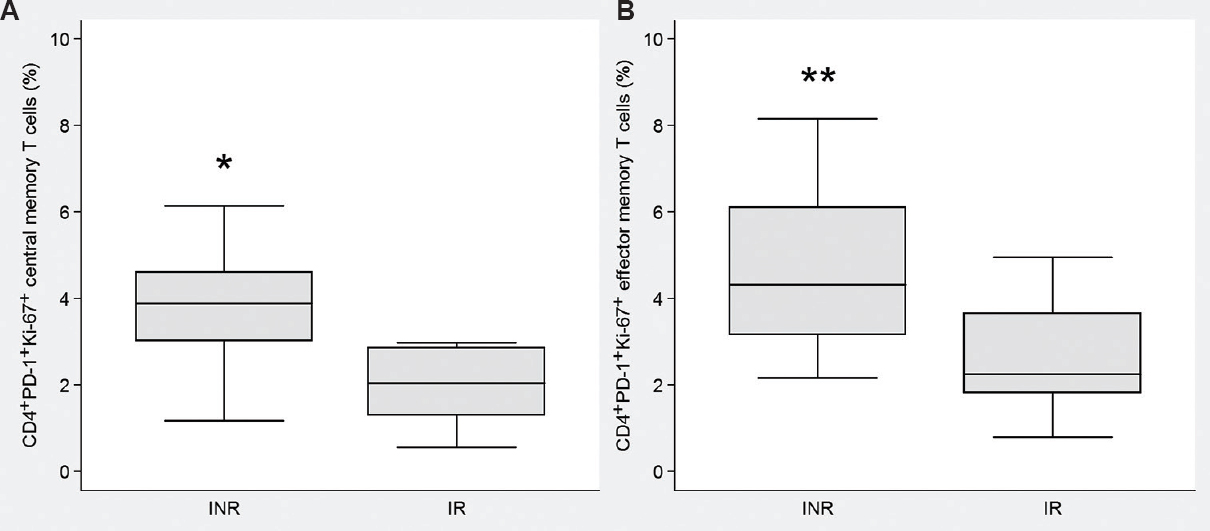
- Frequencies of PD-1+Ki-67+ T cells in INR and IR groups. (A) Central memory subset. (B) Effector memory subset. Medians and quartile ranges are shown. INR, immunological non-responders (n=16); IR, immunological responders (n=21). *P<0.05; **<0.01 compared to IR.
Discussion
In the INR patients suffering from the immunodeficiency, CM and EM CD4+ T-cells proliferated much more extensively than CD4+ T-lymphocytes of individuals responding to HAART and thus having close to normal CD4+ T-cell counts. The number of dividing cells was inversely correlated with the CD4+ absolute counts in the appropriate T-cell subset. As no correlations between the CD4+ Ki-67+ counts and T-cell activation or concentration of proinflammatory cytokines were found in any cell subset, this cell division was probably due to homeostatic proliferation intended to restore the CD4+ T-cell number in lymphopenic hosts20.
It is widely accepted that the intensive cell proliferation leads to the shortening of telomeres at the end of the DNA strands (Hayflick limit)21. This process is the basis for the replicative cell senescence9. Such senescent T-lymphocytes express CD57 molecule on their surface and are capable of rapid interferon-gamma (IFNγ) secretion after antigen stimulation but cannot proliferate in response to the antigen, or produce significant amounts of IL-2, while being prone to spontaneous and activation-induced apoptosis1118. Similar to other groups1822, in our study it was noted that HIV-infection led to unremitting cycles of proliferation and consequent formation of the cell pool enriched with CD57+ ‘senescent’ T-lymphocytes. As T-cell division rates were shown to be higher in the INR patients when compared with the IR patients it was expected to see the growth of the senescent T-cell pool in patients not responding to the therapy. However, in our study no difference was found in the proportions of CD57+ senescent CD4+ T-lymphocytes between the INR and the IR groups. Thus, this study showed that the intensive homeostatic proliferation in lymphopenic HIV-infected patients with the discordant immunological response to the therapy was not associated with the replicative senescence of the CD4+ T-cell pool.
It was also found that the percentage of PD-1+ ‘exhausted’ CD4+ T-cells was significantly greater in the INR patients than in their immunologically responding counterparts. The difference between the groups was most pronounced in the CM but not in the EM group, indicating that the expansion of the EM cells was not the reason for the results received. The true cause for the PD-1 up-regulation in INRs was not clear, but it was suspected that homeostatic proliferation could be one of the factors that was associated with the T-cell exhaustion. It was noted that in the HAART-treated HIV-infected patients the percentage of dividing CM CD4+ T-cells was correlated with the relative counts of the same subsets’ PD-1-positive T-cells, confirming our hypothesis about the role of homeostatic proliferation in the T-cell exhaustion. A different mechanism was probably involved in the CD4+ EM T-cell homeostasis as no correlation was observed between the numbers of cycling and exhausted cells in that subset. The reason for different exhaustion patterns in central and effector memory T-cell subsets is not clear and demands further exploration. Although PD-1 is a distinctive marker of T-cell functional exhaustion16, additional studies using a wider range of exhaustion molecules such as T-cell immunoglobulin and mucin-domain-containing-3 (Tim-3) or cytotoxic T-lymphocyte antigen-4 should be studied to confirm these conclusions. T-cell function (cytokine production and proliferation) should also be assessed in cultures to explore the influence of homeostatic proliferation on the T-cell functional exhaustion.
Several groups have noted that PD-1 is upregulated on the T-cells of HIV-infected patients with discordant response to HAART1622, but the reason for this phenomenon has remained obscure. It is widely accepted that T-lymphocyte exhaustion is induced by the long-term antigen persistence and repetitive stimulation through the T-cell receptor2324 however, our patients were characterized by the high rates of CD4+ PD-1+ T-cells despite their HIV viral load being suppressed. Lin et al25 proposed the idea that in mice constant homeostatic T-cell division could be a reason for exhaustion. These authors showed that after the adoptive transfer of carboxyfluorescein succinimidyl ester-labelled lymphocytic choriomeningitis virus-specific spleen leucocytes into T-cell deficient hosts CD8+ T-cells undergoing acute homeostatic proliferation expressed high levels of PD-1 messenger RNA7. However, no association was noted between the T-cell exhaustion and homeostatic proliferation in humans. Our data on simultaneous expression of Ki-67 and PD-1 on T-cells received from HIV-infected patients with varying efficacy of immunological response to the therapy showed that the proportion of memory CD4+ T-cells simultaneously expressing proliferation and exhaustion markers in non-responders was significantly greater than in the standard response group prompting suggestions that homeostatic proliferation could be somehow connected to the functional T-cell exhaustion.
Acknowledgment
Authors thank Drs M.M. Lederman, D.D. Anthony, and colleagues from the Case Western Reserve University, Cleveland, Ohio, USA for their help in conducting the research.
Financial support & sponsorship: The work was financially supported by the Russian Science Foundation (15-15-0016), Russia
Conflicts of Interest: None.
References
- The stable and permanent expansion of functional T lymphocytes in athymic nude rats after a single injection of mature T cells. J Immunol. 1987;139:1379-84.
- [Google Scholar]
- Peripheral T lymphocytes: Expansion potential and homeostatic regulation of pool sizes and CD4/CD8 ratios in vivo . Eur J Immunol. 1989;19:905-11.
- [Google Scholar]
- Restoration of T-cell homeostasis after T-cell depletion. Semin Immunol. 1997;9:339-46.
- [Google Scholar]
- The peptide ligands mediating positive selection in the thymus control T cell survival and homeostatic proliferation in the periphery. Immunity. 1999;11:173-81.
- [Google Scholar]
- Homeostatic expansion and phenotypic conversion of naïve T cells in response to self peptide/MHC ligands. Proc Natl Acad Sci U S A. 1999;96:13306-11.
- [Google Scholar]
- Incomplete CD4 T cell recovery in HIV-1 infection after 12 months of highly active antiretroviral therapy is associated with ongoing increased CD4 T cell activation and turnover. J Acquir Immune Defic Syndr. 2003;33:125-33.
- [Google Scholar]
- CD4 T cell depletion is linked directly to immune activation in the pathogenesis of HIV-1 and HIV-2 but only indirectly to the viral load. J Immunol. 2002;169:3400-6.
- [Google Scholar]
- Comparative analysis of T-cell turnover and homeostatic parameters in HIV-infected patients with discordant immune-virological responses to HAART. AIDS. 2006;20:1727-36.
- [Google Scholar]
- Replicative senescence of T cells: Does the Hayflick Limit lead to immune exhaustion? Immunol Today. 1997;18:450-4.
- [Google Scholar]
- Reinvigorating exhausted HIV-specific T cells via PD-1-PD-1 ligand blockade. J Exp Med. 2006;203:2223-7.
- [Google Scholar]
- Functional and phenotypic characterization of CD57+CD4+ T cells and their association with HIV-1-induced T cell dysfunction. J Immunol. 2005;175:8415-23.
- [Google Scholar]
- Progressive loss of memory T cell potential and commitment to exhaustion during chronic viral infection. J Virol. 2012;86:8161-70.
- [Google Scholar]
- Molecular signatures of T-cell inhibition in HIV-1 infection. Retrovirology. 2013;10:31.
- [Google Scholar]
- PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature. 2006;443:350-4.
- [Google Scholar]
- Identification of PD-1 as a unique marker for failing immune reconstitution in HIV-1-infected patients on treatment. J Acquir Immune Defic Syndr. 2011;56:118-24.
- [Google Scholar]
- Role of PD-1 co-inhibitory pathway in HIV infection and potential therapeutic options. Retrovirology. 2015;12:14.
- [Google Scholar]
- Immune activation and CD8+ T-cell differentiation towards senescence in HIV-1 infection. PLoS Biol. 2004;2:E20.
- [Google Scholar]
- Discordant responses to cART in HIV-1 patients in the era of high potency antiretroviral drugs: Clinical evaluation, classification, management prospects. Expert Rev Anti Infect Ther. 2016;14:29-40.
- [Google Scholar]
- The peripheral T-cell pool: Regulation by non-antigen induced proliferation? Semin Immunol. 1997;9:347-53.
- [Google Scholar]
- A computational model for telomere-dependent cell-replicative aging. Biosystems. 2008;91:262-7.
- [Google Scholar]
- High T-cell immune activation and immune exhaustion among individuals with suboptimal CD4 recovery after 4 years of antiretroviral therapy in an African cohort. BMC Infect Dis. 2011;11:43.
- [Google Scholar]
- Expression of the PD-1 antigen on the surface of stimulated mouse T and B lymphocytes. Int Immunol. 1996;8:765-72.
- [Google Scholar]
- Expression of programmed death 1 ligands by murine T cells and APC. J Immunol. 2002;169:5538-45.
- [Google Scholar]
- Programmed death-1 (PD-1) defines a transient and dysfunctional oligoclonal T cell population in acute homeostatic proliferation. J Exp Med. 2007;204:2321-33.
- [Google Scholar]




