Translate this page into:
Implication of microsatellite instability in human gastric cancers
Reprint requests: Dr. Prakash C. Sharma, Professor, University School of Biotechnology, Guru Gobind Singh Indraprastha University Dwarka, Sec 16C, New Delhi 110 075, India e-mail: prof.pcsharma@gmail.com
-
Accepted: ,
This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-Share Alike 3.0 Unported, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This article was originally published by Medknow Publications & Media Pvt Ltd and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Microsatellite instability, one of the phenomena implicated in gastric cancer, is mainly associated with the expansion or contraction of microsatellite sequences due to replication errors caused most frequently by mutations in the mismatch repair (MMR) and tumour suppressor genes. Tumours exhibiting microsatellite instability are proven to have truncated products resulting from frequent mutations in mononucleotide or dinucleotide runs in coding and non-coding regions of the targeted genes. Epigenetic changes like hypermethylation of the promoter region of MMR genes as well as gene silencing are also responsible for the microsatellite instability phenotypes. Assessing microsatellite instability in tumours has proved to be an efficient tool for the prognosis of various cancers including colorectal and gastric cancers. Such tumours are characterized by distinct clinicopathological profiles. Biotic agents like Epstein Barr Virus and H. pylori along with other factors like family history, diet and geographical location also play an important role in the onset of gastric carcinogenesis. Instability of mitochondrial DNA has also been investigated and claimed to be involved in the occurrence of gastric cancers in humans. Development of simplified but robust and reproducible microsatellite instability based molecular tools promises efficient prognostic assessment of gastric tumours.
Keywords
Gastric cancer
microsatellite instability
mismatch repair
MSI-H phenotype
mtMSI
tumour suppression

Introduction
During the lifetime of an organism, various cellular processes work as an organisation and are responsible for the overall health status of the organism. Any deterioration or malfunctioning of these processes could induce aberrations in genome, transcriptome or proteome. Such alterations may switch-on the proto-oncogenes and finally develop cancer.
One of the leading causes of cancer associated deaths in the world is gastric cancer (GC), though its incidence has decreased in the last decade1. Prognostic methods applied to detect GC are poor with limited use and pose a major clinical limitation in detecting cancer at an early stage such that less than 5 per cent people survive for more than five years2. In recent times, researchers around the world have reported various distinct GC specific clinicopathological profiles, facilitating cancer prognosis and detection. GC is now generally considered as the outcome of irregularities in complex biological processes involving many genes which regulate activities such as cell growth, death or apoptosis, DNA repair, etc. (Fig. 1). The alterations in gene regulation activities result from various underlying genetic instabilities and epigenetic changes.
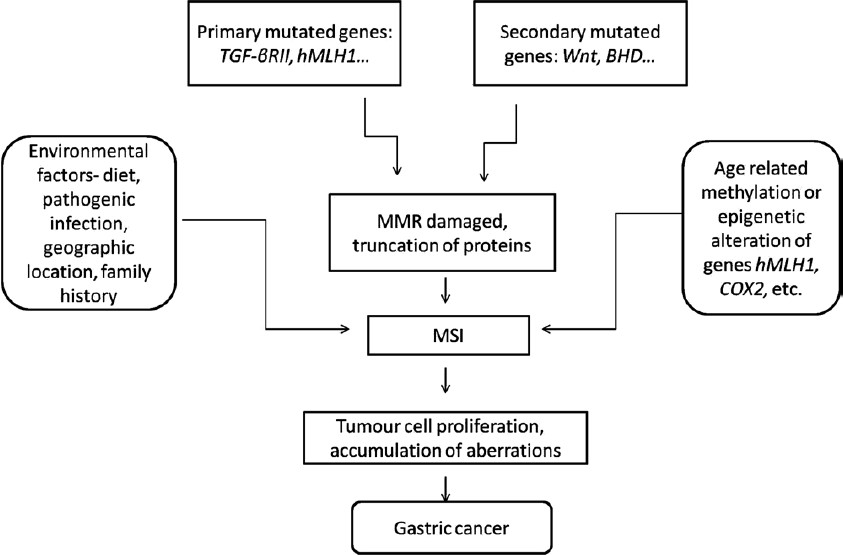
- The Mutator pathway responsible for gastric cancer. Various factors and genes involved are shown.
Once considered as junk DNA, the repetitive elements are now believed to have a significant role in the normal functioning of the cells. The presence of repetitive rudiments in coding and non-coding DNA makes them a valuable region both structurally as well as functionally. The repetitive DNAs are classified into satellites, minisatellites, and microsatellites on the basis of length of repeat units. Microsatellites are short iterations of 1-6 nucleotide long units, non-randomly distributed in both prokaryotic and eukaryotic genomes. Microsatellites are further classified into perfect, interrupted and compound microsatellites (Fig. 2). During the past decade microsatellites have emerged as molecular markers of choice for diverse applications owing to some advantages over the other marker systems. Microsatellites undergo mutations at a very high rate ranging from 10-6 to 10-2 per generation3 and thus, are highly polymorphic in nature. The disorders in the microsatellite regions like insertions, deletions, etc. may result in altered expression of associated genes finally changing the phenotype of the organism. These genomic alterations are named microsatellite instability and are now considered as markers for prognosis and diagnosis in many types of cancers.
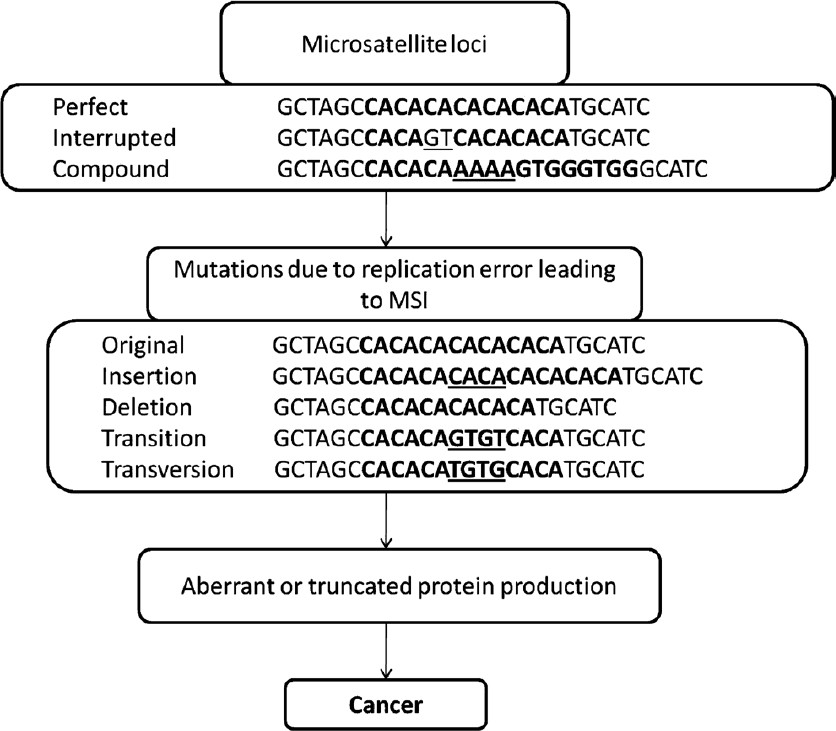
- Types of microsatellites and different aberrations involved in the incidence of cancer. MSI, microsatellite instability.
The genomic instability pathways mentioned in the literature till date are of two types: chromosomal instability (suppressor pathway) and microsatellite instability (mutator pathway)4. The former includes tumour suppressor gene inactivation commonly caused by mutation or allelic loss. Loss of heterozygosity (LOH) is thought to contribute to tumour suppressor gene inactivation and has been detected in many types of human tumours. Genomic locations demonstrating high rates of LOH represent loci that potentially anchor tumour suppressor genes. The alteration in the microsatellite DNA due to polymerase slippage results in microsatellite instability (MSI). Accordingly, these two mechanisms are related to different GC subtypes such as intestinal type associated with MSI and diffuse type linked with suppressor phenotype56.
Cancer can be diagnosed at an early stage by assigning MSI status to the cancerous tumour or cells. In last few decades, a large amount of data have been accumulated on various genes, mechanisms, features and agents responsible for causing gastric cancer. This review covers various factors and features of microsatellite instabilities- mitochondrial or nuclear, associated with gastric cancer.
Origin and detection of microsatellite instability
Microsatellites are considered hypervariable and thus contribute towards species and population diversity. The mutation rates at microsatellite loci differ with regard to repeat unit length (mono, di, tri, etc.), microsatellite type (perfect, compound, etc.), base composition and taxonomic groups67. Microsatellite flanking sequences have the ability of modifying their genomic context as well as mutability of the locus89. The hypervariability in the microsatellite tracts arising due to DNA polymerase slippage following re-alignment of nascent and template strands and if this alignment remains unobstructed, then repeat number is altered610. Mismatch repair (MMR) system corrects these alterations and reduces the error rate by 100-1000 fold, together with DNA exonuclease proof reading ability1112. Some MMR genes themselves contain microsatellites in the coding regions. Therefore, if the MMR gene itself gets mutated, then the repair of the mutated region elsewhere in the genome is hampered. Moreover, the incidence of carcinogenesis increases with the increase in mutation rates in the MMR genes.
Different methods have been employed to study the prevalence of MSI in GC over the last decade. Polymerase chain reaction (PCR) is the most common technique used in the prognosis of MSI and classifying tumours on the basis of MSI status. PCR amplification has been used to study expression analysis of MSI by comparing PCR profiles obtained from both normal and cancerous tissue13. One major improvement in MSI detection is multiplex PCR amplification. The advantage of multiplex PCR over the normal single locus PCR is that more than one microsatellite loci under investigation are amplified within one reaction, decreasing the number of steps involved as well as maintenance of data in simplistic way. For obvious reasons, the fluorescent analysis has an upper hand over the radioactive detection system.
Association of MSI with cancer
MSI plays a very important role in the development of cancer14. In 1993, MSI was implicated as one of the factors responsible for the occurrence of colon cancer designated as hereditary nonpolyposis colon cancer (HNPCC)15. Soon after, several studies suggested the role of MSI in other cancers also like endometrial and gastric cancer1617. In 1997, a meeting held at Bethesda, USA, during National Cancer Institute Workshop proposed a panel of five markers including mononucleotide (BAT26 and BAT25) and dinucleotide (D2S123, D5S346 and D17S230) markers for the uniform detection of MSI tumours18. Accordingly, the tumours were classified into three classes based upon the MSI status: tumours with instability at more than two loci designated as MSI-High (MSI-H), those with instability at two loci as MSI-Low (MSI-L), and those which do not show instability at any of the microsatellite loci as MSS tumours. Research groups have also classified human cancers on the basis of microsatellite length alterations as Type A (< 6 bp length change) and Type B (>8 bp length change)19. Various genes involved in MMR machinery are reported to undergo mutations as well as hypermethylation, resulting in the truncation of the encoded protein product of the respective gene. The mutations in these genes are responsible for the MSI phenotype resulting in cancer progression in the affected cell lines2021. Furthermore, diverse populations of the world have different MSI prominence due to various other environmental factors ensuing dissimilar outcomes. MSI has been reported in around 5 to 50 per cent of sporadic GC22–24.
Target genes and mutations involved
MSI is responsible for aberrations in several genes involved in normal functioning of the cells. These mutations lead to the truncation of the gene products and/or suppression of the gene activity. Over a decade, different genes have been discerned to undergo mutations at the repetitive sites viz. tumour growth factor beta receptor type II (TGF-βRII), Bcl-2 associated X protein (BAX), hLMHI, hMSH3, hMSH2, hMSH6 and insulin growth factor type-II receptor (IGF-IIR) genes25–2741 (Table). These mutations are not only confined to intronic regions of the genes but also appear in the coding regions. Moreover, epigenetic changes like hypermethylation has also been implicated in various studies, for example, hypermethylation of hMLHI, RAB32, CDH1, etc. Several targets of MSI are enlisted below which undergo expansion, contraction, point mutation and rearrangements in the microsatellite region.
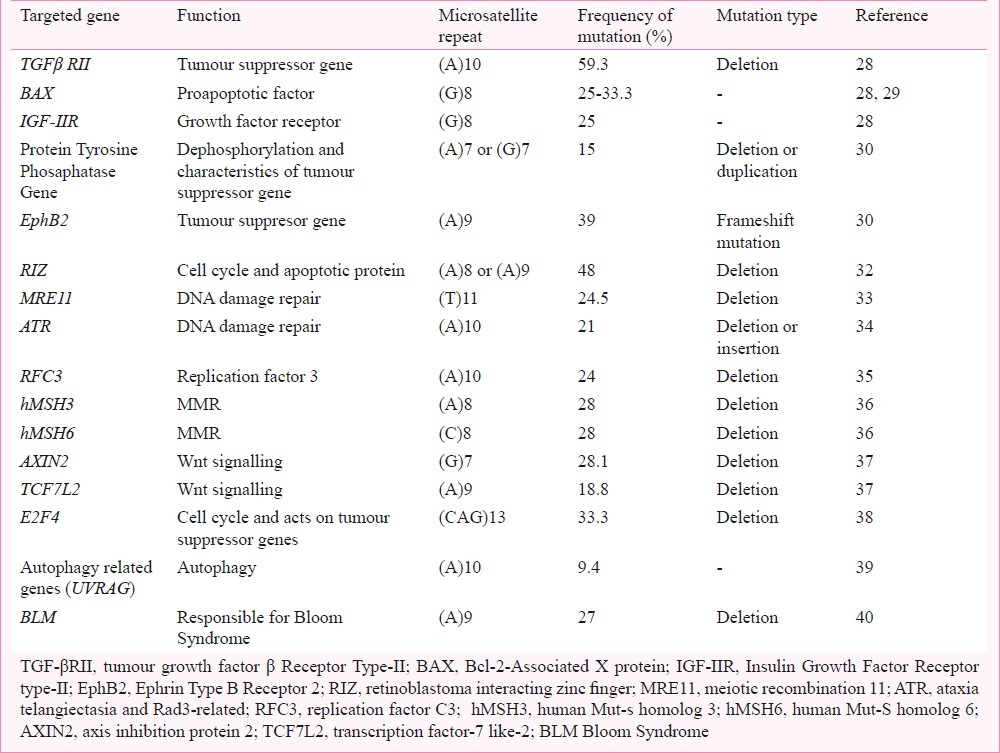
Tumour suppressor genes
During 1970s and early 1980s, evidences that suggested the involvement of a different class of genes other than proto-oncogenes in cancer started accumulating. These genes have the property of suppressing the growth of abnormal cells by interfering with their cellular machinery. Later, these were named as tumour suppressor genes and alterations in these genes result in the altered phenotype and finally, cancer. The tumour suppressor genes can undergo mutations either by suppressor pathway or by mutator pathway. p53 is the most studied tumour suppressor gene that follows the suppressor pathway resulting in the cancer progression. Other genes like TGF-βRII, gene coding for tyrosine phosphatase, protein kinases coding gene, EphB2, and retinoblastoma protein-interacting zinc finger (RIZ) gene follow the MSI pathway4243. These genes are affected due to frameshift mutations in the microsatellites spanning the coding region causing the loss of function and ultimately leading to failure of translation. The loss of function of TGF-βRII is considered to be the first step in the onset of cancer. Reports have shown frameshift mutations in TGF-βRII in 59.3 per cent of MSI-H associated GC28. Mutations at this locus are supposed to be the primary target for alteration and aberration in the normal phenotypes. Association of TGF-βRII with intestinal type or glandular structure has been reported in GC with a better survival rate44. During gastric tumorigenesis, TGF-βRII mutations play pivotal role such that GC progresses by escaping the growth control signal of TGF-βRII network45.
Alterations in dual specificity phosphatase (DPTP) associated with MSI-H are reported to be around 12 per cent (1 bp deletion) and thus, contribute to development of cancer along with other primary mutations30. DNA-dependent protein kinase (DNA-PKC) genes have two mononucleotide repeats poly(A)8and poly(A)10, of which, latter exhibits frameshift mutations4647. The frameshift mutations in this tract follow an expression loss mechanism and are found associated with lymph node metastasis and neutrophillic infiltration in GC48.
Inactivation of APC (adenomatous polyposis coli) gene plays a role in the development of GC. Similar to that in CRC, APC gene undergoes mutations during the early developmental stages of GC49. Mutations of APC send a downstream signal resulting in further mutations in EphB2 gene that belongs to the family of receptor tyrosine kinases and has been extensively studied because of its emergence as a tumour suppressor gene in CRC43. A high incidence (39%) of EphB2 mutation in (A)9 tract occurs in GC-MSI cases whereas no significant relationship with clinicopathological features has been reported31. EphB2 has a role in the developmental processes particularly in the nervous and vasculature system. The presence of EphB2 mutations in stomach mucosa results in gastrointestinal cancer50. While this gene is a target in mutator pathway, it is mutated in endometrial cancer by following a different tumorogenic pathway31.
RIZ, a member of tumour suppressor genes, coding two proteins RIZ1 and RIZ2, is involved in chromatin mediated gene activation and silencing. RIZ1 (PR+) product is considered as a tumour suppressor candidate on region 1p36, which is found deleted in most types of human cancers. In MSI(+) tumours, RIZ is affected by frequent frameshift mutations in one or two coding poly(A) tract, an (A)8 tract at the coding nucleotide sequence 4273-4280 and an (A)9 tract at 4462-4471 in exon 832. A literature survey showed 48 per cent of RIZ1 mutations associated with GC, 33 per cent with endometrial cancer and 26 per cent with CRC32. During tumorigenesis, biallelic mutations of RIZ are proposed to be clonally selected that have a more important role in endometrial cancer over GC and CRC32. The RIZ gene is mostly affected as high as in 57 per cent of cases in the poly(A)9 (mostly deletion) tract in MSI-H tumours over MSI-L and MSS56. It can be said that RIZ mutations may have a role in GC in MSI-H and provide an important mutational target in GC as in the case of endometrial cancer.
Tumour suppressor genes are known to be involved in the onset of gastric cancer. Mainly the TGF-βRII is one of the primary genes which has a direct consequence, as also reported in CRC. The prevalence of LOH in these genes is less in comparison to MSI and, therefore, one can suggest the involvement of mutator pathway. The preference for the MSI pathway may be the outcome of the presence of mononucleotide repeats in the coding sequence of these genes. Such repeats have a biasness to undergo mutations at a higher rate over other repeats and thus follow the mutator pathway over the suppressor pathway52.
Mismatch repair and DNA damage repair genes
Mismatch repair (MMR) system is known to be responsible for correction of any error arising during DNA replication. To maintain the genomic fidelity, the MMR system has to be efficient in correcting these mutations. MMR system genes like hMSH2, hMSH3 and hMSH6 play a pivotal role in correcting these errors53. Gene hMSH2 forms a heterodimer with hMSH3 or hMSH6 and binds with the part of DNA harbouring the error. Other genes which take part in this process are MRE-11, replication factor C3 (RFC-3) and checkpoint genes Ataxia telangiectasia and Rad3-related (ATR) and CHK133–35. All these genes contain mononucleotide repeats that undergo alterations.
hMSH3, hMSH2 and hMSH6 are homologs of mut-s genes present in bacteria. Biallelic and monoallelic mutations are reported at hMSH3 and hMSH6 loci exhibiting MSI phenotype36. Various types of mutations including frameshift or indels in the mononucleotide repeat tracts have been observed in these genes resulting in either loss of function or low expression of the genes involved in GC. Low expression of hMSH2 gene was reported in moderately and poorly differentiated gastric cancers showing its metastasis and prognostic significance36. A significant association between MSI-H phenotype and MRE-11 mutations (intronic) has been suggested to be a novel target in MSI-H GCs33. MRE-11 gene, one of the novel targets in GC, is involved in the progression of GC at later stages.
Ataxia telangiectasia and Rad3-related (ATR), a DNA damage repair gene, is vulnerable to somatic mutations that normally occur in sporadic MSI positive GC tumours34. In association with CHK1, it induces cellular check in G2-M phase through the inhibition of Cdc25c and Cdc2 by phosphorylating these two proteins. The hotspot of these mutations is a short stretch of (A)10 repeat. Insertion or deletion of nucleotides generates (A)11 or (A)9 repeats consequently resulting in cancer phenotype. These reports suggest that ATR, and CHK1 are some of the direct targets of the mutator pathway in stomach tumorigenesis. In addition, the inhibitory action on the pathway of ATR-CHK1 DNA damage-response could result in the tumorigenesis of GC with MSI.
RFC3 and PCNA (proliferating cell nuclear antigen) help in the process of proofreading of DNA. Recent studies have revealed the presence of (A)10 repeat in exon 3 of RFC3 gene and an (A)7 repeat in exon 13 of RFC1 gene which can provide a potential mutation target in cancer with MSI35. Association between MSI-H and RFC3 mutation has been recorded in around 40 to 69 per cent of GC, and more frequently in CRC, without any significant relationship with clinicopathological features35.
Wnt signalling pathway genes and transcription factors
Wnt (Wingless-int) signalling pathway is involved in the regulation of morphogenetic events during development, for example, gut development. The Wnt genes, TcF/Lef family transcription factors and APC gene work in a feedback manner. The Wnt genes bind to the frizzled proteins and activate Wnt signalling pathway. This activation inhibits APC/AXIN/GSK3β complex resulting in the release of β-catenin which finally binds to transcription factors TcF/ Lef and translocates into the nucleus54. Mutations in APC and Wnt pathway genes [AXIN2-poly(G) and TCF7L2-poly-(A)] have been reported in different types of cancers36. The Wnt genes contain exonic mononucleotide repeats and are supposed to be tumour suppressors as these are negative regulators of Wnt signalling54–56. Although mutations in (A)9 repeat (deletion-1bp) of TcF-4 gene have been implicated in 14.3 per cent of CRC cases, no single case of GC had mutation in this gene58. In a recent study, it was observed that 28.1 per cent of AXIN2 and 18.8 per cent of TCF7L2 with frameshift mutations in the mononucleotide repeats were associated with the MSI-H cancers, and no single mutation was found in MSI-L/MSS cancers37. Till now, not much is known about how these genes mark the onset of GC and related clinicopathological features. Further investigations are required to explain the possible role of these genes in the incidence of GC.
Other targets of MSI
IGF-IIR gene belongs to the insulin growth receptor family and is thought to be an important gene in the progression of GC. The gene contains (G)8 microsatellite similar to that in BAX gene (an apoptotic gene). BAX alongwith E2F4 having trinucleotide repeats is confirmed to have a role in carcinogenesis of stomach. Investigators have reported that among MSI-H GC, these genes exhibit frameshift mutations causing the loss of expression59. Around 25 to 33 per cent mutations in the coding mononucleotide repeat of IGF-IIR and BAX genes were reported in MSI-H GC28. E2F4 mutations were present in the early stages of multiple GC and exhibited deletions in the microsatellite region suggesting that E2F4 is a mutational target for MMR defects38. It can be said that all these genes in one or the other way follow pathway similar to mutator pathway instead of suppressor pathway as no significant relationship between LOH and the mutations in these genes is reported.
In 1977 Birt-Hogg-Dube (BHD) syndrome was described as a rare form of autosomal dominantly inherited syndrome exhibiting characteristics like fibrofolliculomas, trichodiscomas and acrocordon skin abnormalities60. The genic region contains poly(C)8 mononucleotide repeat in exon 11 and around 44 per cent of BHD patients are reported to have undergone germline mutations61. The repetitive region is a hotspot for indels and frameshift mutations that cause truncation in folliculin protein. Hence, BHD is now considered as a tumour suppressor gene62. In GC with MSI-H, 16 per cent cases of mutations in BHD gene have been reported63. The reports have also shown mutations in BAX and TGF-βRII in BHD mutated GC cases. To sum up, BHD mutations are a rare event in MSI-H GC and occur downstream to BAX and TGF-βRII mutations.
Autophagy (ATG) is a process considered as a type-II programmed cell death (PCD) and has a relationship with apoptosis. The former has a role in cell survival also64. Mutations in the (A)10 repeat of UVRAG (ATG) gene in 9-28 per cent and 18-28 per cent cases of GC and CRC, respectively were reported in MSI-H cases39.
Bloom syndrome (BLM) gene undergoes frameshift mutations, occurring in poly(A)9, resulting in the generation of a truncated and non-functional BLM protein. The aberration at BLM gene is known to cause Bloom syndrome which is a pre-malignant situation characterized by genomic instability and high mutational rates. An inverse relationship of BLM gene with TGF-βRII mutations was reported and the relationship was more evident when considered along RAD50 gene28. Loss of BLM expression by deletion of trinucleotide and mononucleotide repeats due to MSI results in the increase of the genetic irregularity of an already present unbalanced genotype in gastric tumours40. The role of BLM in GC has been proposed to be of a major kind associated with hMSH3/hMSH6 mutation but is a secondary mutator phenotype.
The changes in the function of a gene could be due to genetic or epigenetic changes. The latter do not affect the underlying DNA sequence rather these change the function of the gene by processes like methylation, acetylation, etc. These changes can persist through generations like the germline mutations. In gastric cancers, many genes undergo hypermethylation. hMLH1 gene is one of the most studied genes in the incidences of cancer. Earlier, mutation in coding region of hMLH1 gene was thought to be responsible for the MMR deficient phenotype in GC, but now through several studies the hypermethylation of CpG island region in the promoter of hMLH1 has been found responsible for MSI in GC patients66. For the inactivation of hMLH1 gene, methylation at a small region (from -270 to -199) proximal to transcriptional start site is important and consequently may result in MSI in a subset of GC cell lines. In addition, hMLH1 hypermethylation occurs chiefly in the surroundings of HPP1 (other related gene) hypermethylation. It can be said that HPP1 hypermethylation occurs at early stages of GC in MMR deficient cells. A correlation exists between MSI phenotype and CDH1 promoter methylation postulating that during methylation process, entire group of genes may be jointly methylated. The silencing of these genes by hypermethylation of promoters may participate in carcinogenesis through the microsatellite instability pathway67.
Taken together, the targets of MSI in gastric cancer are mostly harbouring mononucleotide repeats that are generally altered by frameshift mutations or indels (Table). Moreover, the incidence of gene mutations in GC is quite similar to CRC (Fig. 3)21,29323642. These events further lead to change in phenotype governed by the respective genes.
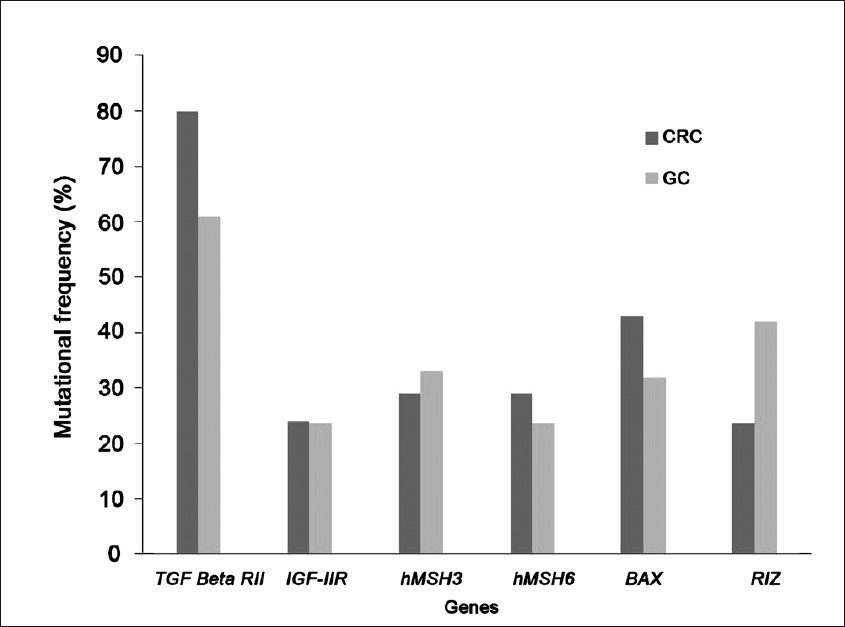
- The frequency of mutation in various targeted genes in sporadic colorectal cancer (CRC) and gastric cancer (GC).
According to the loss of function during the onset of GC, two types of genes have been proposed by Perucho68: (i) Primary mutated genes: these genes are responsible directly for the occurrence of GC. Genes under this category are mutated at first in the molecular pathway. For example, TGF-βRII, BAX, hMLH1, etc. (ii) Secondary mutated genes: genes which are indirectly mutated in the carcinogenesis of the stomach. These are regulated downstream of the primary mutated genes. These have a meandering effect on GC phenotype. These include Wnt genes, BHD gene, tyrosine phosphatase kinase gene, etc.
Accordingly, the genes get activated in early or late phase of the carcinogenesis depending upon the above classification. As switched on and off in a signal transduction pathway, the abnormal primary mutated genes direct the mutation in the secondary target genes. Also, the rate of mutation increases in the secondary mutated genes following aberrations in the primary mutated genes.
Clinicopathological implications of gastric cancer with MSI-H phenotype
Various clinicopathological features are associated with GC which includes GC type- intestinal or diffuse, stage, survival rate, location, mucin type, lymphoma association, age and sex (Fig. 4).
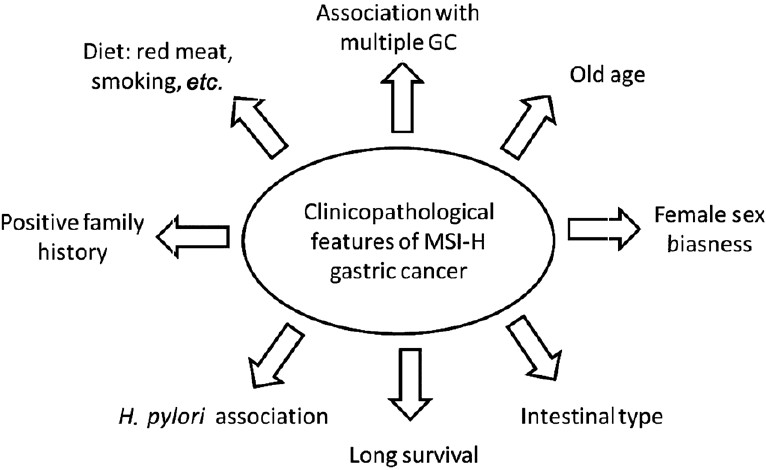
- Various clinicopathological features associated with gastric cancer in MSI-H tumours.
Histological and phenotypic features
As mentioned earlier, GC is divided into two classes: intestinal or diffuse type. The intestinal type is preceded by a process spanning various stages starting from normal mucosa, followed by chronic gastritis, atrophic gastritis, intestinal metaplasia, dysplasia and finally cancer. The first stage of gastritis is due to the deterioration of the normal mucosa succeeded by glandular loss and intrusion of inflammatory cells into the glandular zones in atrophic gastritis. In the next step, the normal mucosa is replaced by intestinal type epithelial cells and at last stage before acquiring the state of cancer, the cells gain the potential to become cancerous and metastasize. Most of the studies till now have shown strong association of MSI-H in GC with intestinal type. Several reports have shown the association of GC with MSI or LOH6970. MSI-H was reported highest in GC and GC-intestinal metaplasia cases when the tumour was extracted from the upper third of stomach whereas, LOH was detected frequently in the cases having lymphatic and vascular invasion in GC and GC-intestinal metaplasia71. Intestinal type GC undergoes much genomic instability in comparison to diffuse type. These reports not only help in better prognosis but also tell about the exclusive features shared by the two types of GC.
MSI-H status also varies with the stage of the GC. MSI-H presence is more prevalent in early phase of the four developmental steps72. MSI caused by hMLH1 methylation has a very important role in GC at stage IV73. The other clinicopathological features associated with GC are antral location, female sex biasness, older age, high survival rate, and low lymph node metastasis74–76.
Age and sex
Young patients (<40 yr of age) around the world, account for only <5 per cent of all GCs77. Comparison of the occurrence of GC in young patients with older patients suggests that the former show a more deleterious clinical course with poor prognosis. Therefore, the genetic profile of younger GC patients might be different from that of older patients and both show different clinic-pathological features. For example, gastric tumours in elderly are generally located in the lower third region, show relatively low metastasis and are present in 8-15 per cent of synchronous GCs whereas the younger patients have tumours in middle third region with relatively high metastasis occurring in 3 per cent synchronous GCs78–80.
Epigenetic changes also play a major role in GC incidences in elderly patients. The methylation of hMLH1 gene and its loss of expression increases with increase in the age of the GC patient81. Age related gene methylation may have an important role in increasing the chances of development of malignant neoplasms in older patients as CpG island methylation is a dominant mechanism for gene inactivation. For example, in elderly people clinicopathological characteristics like poorly differentiated medullary type adenocarcinoma of intestine could be due to an epigenetic event within the hMLH1 gene involving hMLH1 promoter hypermethylation82. A strong positive correlation between MSI and GC in young female over young male patients has been exhibited83. These results suggest involvement of different molecular pathways in the onset of GC in male and female patients such that one of these may follow mutator pathway whereas the other follows suppressor pathway.
Multiple gastric cancers, mucin phenotype and lymphoma development
Once initiated, a cancer can further spread out to surrounding areas resulting in multiple cancer types in different body parts or in the same region. The results published till date convey that either the genetic makeup or the environmental factors of the patients are responsible for the occurrence of the multiple GC with very high MSI6771. The tendency of multiple gastric cancers is also directly proportional with age, for example, old aged persons are more prone to development of multiple GCs73. Studies have shown the usage of MSI as a molecular marker for the prediction of multiple GCs23. MSI not only results in carcinogenesis but also promote the occurrence of multiple GCs over solitary cancer73. Patients having GC with MSI-H show higher prevalence of secondary GC in comparison to patients with MSI-L or MSS GC. Some genes like TGF-βRII, BAX and hMSH3 undergo higher mutations in the type I synchronous carcinomas as compared to type II synchronous carcinomas suggesting that MMR system impairment might have an important role in carcinogenesis67.
One of the basis of classifying gastric carcinoma is the presence of extracellular mucin in tumours (atleast 50% of tumour volume) as defined by World Health Organization84. Mucin type or mucinous gastric cancer (MGC) comprises 2-6 per cent of all GC types85. Its association with MSI and associated clinicopathological features are still debatable. Variable levels of association, from low to high, have been reported between MSI and MGC in different reports3185. Researchers have also tried to correlate the two by comparing MSI and mucin phenotype in multiple GC and solitary GCs. Early multiple GCs and early solitary GCs display different mucin phenotypes. The early multiple GCs had dominant mucin phenotypes as well as MSI frequency86. These results suggest that mucin phenotype alongwith MSI may aid in prognosis of early GCs as compared to advanced GCs.
Mucosa-associated lymphoid tissue (MALT) lymphomas are extranodal low-grade B-cell tumours, developing in the stomach and in other organs also. Genetic instability was recorded in 69 per cent patients with gastric MALT lymphoma, of which 54 per cent displayed replication-error-positive phenotype87. MSI has been speculated to have a direct role in MALT lymphomas, however, convincing evidences are still lacking. For the analysis of MSI, markers neighbouring the chromosomal loci involved in lymphoma should be used to follow ‘Real Common Target Genes’ model. This model entails that a specific group of genes called real target genes, having microsatellite repeats undergo high frequency of mutations as compared to other microsatellite positive bystander genes and assist in tumour growth8889.
Dietary factors, familial connection and demographic biasness
Various factors associated with the occurrence of GC also include diet factor and family history affecting the MSI status. A weak positive relationship between family history (if affected person is mother only) is ascertained with 2 bp deletion in the MRE-11 gene45. High consumption of red meat and meat sauce, nitrite intake, total protein level and sodium intake affect the normal phenotype and transitions to GC in MSI-H lines90. Alcohol consumption, vitamin C and cigarette smoking induces GC with hypermethylation of hMLH191. The exact mechanism by which alcohol consumption and cigarette smoking leads to GC is not known.
Majority of GCs belong to sporadic type and of these only 10 per cent accounts for familial aggregation. Germline alterations in CDH1 gene has been identified in families with clustering of early onset diffuse GC known as hereditary diffuse GC. The pattern involves lower frequency of CDH1 mutation (20%) in families from countries with high incidence of GC and countries having lower prevalence of GC with high CDH1 alterations (50%)92. Family having a case history of GC is likely to have progeny exhibiting GC phenotype. People taking animal protein rich diet have adverse effects on GC prognosis among the positive family history cases90. Remarkably, the difference in sporadic and familial cases was only with regard to the age of onset and gender while sharing other clinicopathological features. MSI is strongly correlated to familial GC in contrast to HNPCC93.
It has been noted that populations inhabiting different geographical regions in the world have variable GC status with higher cases in Asia as compared to USA and Europe1. The difference in population response to GC subtypes is due to environmental factors, diet, genetic predisposition and association with Helicobacter pylori. Korean and American patients did not show any marked difference between MSI-H and MSI-L status22. Others have reported a highly susceptible Italian population towards GC where genetic alterations in the non-invasive neoplasia are due to MSI94. In European populations with a very high risk of GC, alterations of MMR system are thought to be prevalent during the early molecular events in carcinogenesis of stomach. MSI prevalence in Japanese GC samples was higher in comparison to samples from American patients of European descent and same pattern was seen for advanced tumour cases among Japanese patients and American patients95.
MSI detection for chemosensitivity
A chemotherapy regime, neoadjuvant chemotherapy, based on 5-fluorouracil (FU) and cisplastin is frequently used to treat advanced gastric carcinoma. The most important aspect in the treatment of neoadjuvant therapy is the precise knowledge of individual's response to the treatment that depends on the genetic makeup as well as different genetic alterations in the cells. Various studies have reported the prognostic exploitation of MSI status in chemotherapy. MSI-H phenotype did not play an important role in predicting any benefit of neoadjuvant chemotherapy on overall survival in GC and has no correlation with chemosensitivity as proved by an in vitro sensitivity test96. Other reports have shown a major difference between chromosomal instability and MSI with regard to response to neoadjuvant cisplastin based chemotherapy. Resistance of cell lines to chemotherapy due to apoptosis escape (loss of p53 gene or damaged MMR system) and the importance of p53 mutations and MSI for predicting the response to neoadjuvant FP chemotherapy in gastric carcinoma have been reported97.
Pathogenic agents responsible for MSI in gastric cancer
One of the causative factors of GC in humans includes involvement of pathogenic agents. The two most studied ones are Epstein Barr Virus (EBV) and Helicobacter pylori. These factors involve a mechanism which results in MSI of the MMR system and finally the cancer phenotype. Some of the facts regarding these two pathogenic agents are mentioned below.
Epstein Barr virus
The EBV is an omnipresent human virus causing several human malignancies. At least, 10 per cent cases of GC are due to the pathogenesis of EBV in the stomach98. The mechanism of carcinogenesis through EBV remains unclear. Epigenetic changes like methylation of the CpG islands of promoter region of the genes like p16, and hMLH1 are common in EBV associated GC99. Methylation of CDH1 gene has invariably been recorded in EBV associated GC but its correlation with MSI was found to be significant66. Experiments with de novo carcinomas elucidated a mutually elite pattern between the presence of EBV and MSI positivity that are independent of each other100. These results convey that MSI in GC and EBV infection of the GC involve different molecular pathways of carcinogenesis.
Association of EBV with lymphoepithelioma like carcinoma or medullary carcinoma of the stomach is a rare type of gastric carcinoma and is described as tumours with histological similarity to nasopharyngeal carcinoma. This state is called as gastric carcinoma with lymphoid stroma101. Two subsets of GC with increased number of lymphocytes are classified as EBV positive cancers and MSI-H cancers. The CD3 (+) and CD8 (+) tumour infiltrating lymphocytes are characteristic of MSI and MSS/EBV (+) associated GCs which can be used as favourable prognostic factor, independent of the pathogenesis of GCs102. However, other workers have reported no beneficial role of EBV as a prognostic factor in lymphoepithelioma GC over MSI103.
Helicobacter pylori
The Gram-negative microaerophilic bacterium H. pylori inhabits stomach of at least half of the world's population101. Exposure to the bacterium in the childhood can prolong the infection for the rest of the life of the host if not diagnosed and treated at right time. The prolonged contact with the bacteria results in the carcinoma of the stomach and is considered as a class-I carcinogen. The risk of developing GC is related to the heterogeneity of H. pylori virulence factors, namely the Cags pathogenicity island and the vacuolating cytotoxin VacA105.
H. pylori infection has a negative effect on the MMR system and the activity of various MMR proteins like hMLH1, PMS1, PMS2, hMSH2 and hMSH6 gets significantly diminished106. The decrease in the expression of the genes is dose dependent and independent of the virulence factor CagA. The cells, in which H. pylori has been eradicated, return to normal levels of hMLH1 and hMSH2 proteins signifying a reversible inhibition of MMR gene expression. Other reports of methylation of CDH1 in cases with H. pylori-CagA+ phenotype comparative to H. pylori-CagA- ones in intestinal type GC are also available107. An inverse relationship between MSI and CagA protein has also been reported suggesting that other factors are also responsible for MSI in GC in addition to the bacterium CagA protein107. GFP reporter-based in vitro assay demonstrated that H. pylori infection induces MSI, linked with low expression of the MMR proteins hMLH1 and hMSH2108. H. pylori induces genomic instability of (CA) n repeats in mice resulting in impairment of MMR machinery and generating a transient mutator phenotype making the gastric epithelia susceptible to aggregation of genetic instability leading to gastric carcinogenesis109. A recent review110 on H. pylori associated GC has proposed a model to explain how the bacterium causes carcinogenesis. Three steps have been proposed: increase in DNA damage frequency and decrease in repair activity, mutations of mtDNA and finally, induction of a transient mutator phenotype upon infection with H. pylori110. While some researchers contradict above findings and suggest that both H. pylori negative and positive tumours showed same amount of MSI in GC and even after eradication of the bacteria there were no changes in chromosomal aberrations111 thereby suggesting that H. pylori infection act as a synergistic factor in GC but not a direct factor causing carcinogenesis by altering the gene expression. With the progression of gastric lesions, the methylation of repetitive elements like SINEs, LINEs and satellites increases regardless of the H. pylori infection. To sum up, most reports suggest that H. pylori is a leading factor for causing GC by damaging the MMR machinery.
Mitochondrial microsatellite instability
The development of GC is a complex process during which a large number of mutations arise in nuclear and mitochondrial DNA (mtDNA). Human mtDNA is a circular genome composed of 16569 bp and encodes 13 polypeptides of mitochondrial respiratory chain, 22 tRNA and 2 rRNA required for protein synthesis112. Several repetitive elements like mono- and di-nucleotide repeats are present in the mitochondrial genome. Of these, the most suitable region for studying mitochondrial MSI (mtMSI) is located in the D loop region. Two important sites include a (CA)n microsatellite repeat starting at 514 bp and a homopolymeric C tract present between the nucleotide bases 16184 and 16193 bp which could be interrupted by T at 16189 bp113114. The former region contains some regulatory sequences which are important for the normal functioning of the cells. Mitochondrial genome is susceptible to around 10-100x mutations because of its structure and the nature of replication machinery115. The mt genome is also vulnerable to oxidative damage due to high reactive oxygen species (ROS) concentration in the vicinity of the organelle alongwith the poor MMR machinery. Other than CRC, mtMSI has also been reported in case of gastric cancer116. Two components, ROS and defective MMR, are responsible for the mtMSI in H. pylori-associated GC117. Various mutations in the D loop region in GC phenotype reflecting insertions, deletions, transitions and frameshifts were encountered. Some genes like ND1, ND2 and ND5 (subunit of NADH dehydrogenase) provide a hub of mitochondrial genetic instability involved in gastric dysplasia and GC.
The clinicopathological characteristics of mtMSI+ gastric cancers remain unclear. No obvious relationships between mtMSI and tumour size, depth of invasion, node metastasis or clinical stages were detected indicating a limited role of mtMSI in predicting the prognosis of gastric carcinomas. Insertions as well as deletions in the D-loop region of the mtDNA and transitions in genes like ND1, ND5 and CO1, were found in GC samples but have no association with MSI118. It seems that as the carcinogenesis progresses, the level of mtMSI also elevates and thus, mtMSI has a significant function in the onset of GC. In tumoral cell mtDNA, a ~8.9 Kb deletion is more prevalent as compared to other mutations. This mutation is also related to a particular age group (40-50 yr) and intestinal type of GC119. mtMSI is an early and valued event in the succession of GC, that too of intestinal type. ROS, genetic irregularity, environmental factors and poor efficiency of mtDNA repair machinery cause such deletions. Suggestingly, mtMSI can be used as a prognostic marker for GC prediction at a particular state. Moreover, the use of mtMSI as a prognostic marker aid in the identification of high-risk dysplasia that may develop into intestinal type GC120.
mtDNA mutations are associated with H. pylori infection causing chronic gastritis and peptic ulcer tissues indicating that the consequences of H. pylori infection are the aggregation of mutations in mtDNA at early phases of GC development109121. The bacterial infection directed high frequency of mutations in the D-loop region alongwith genes ND1 and CO1 of mtDNA of gastric cells109.
Some tumour suppressor genes are reported to be associated with the instability of mitochondria of which RUNX3 is the one recently reported. RUNX3 belongs to runt related transcription factors (RUNXs) and undergoes methylation producing the ineffective RUNX3 protein122123. In several studies, the MSI-H, mtMSI and RUNX3 promoter methylation implicated in GC have been associated with several clinicopathological variables, although different reports lead to different conclusions118124–126. No association with any of the clinicopathologic variables are reported whereas mitochondrial instability only proved to be associated with the tumour node metastasis127. mtMSI and nuclear MSI-H GC evolution is resultant of methylation of RUNX3 gene as suggested by these events127.
mtMSI is a new field for investigation as a causative agent for development of cancer. Recent reports have shown its association with several mutations and finally with carcinogenesis whether it is colorectal, gastric or female cancer. The D-loop region of the mitochondria is highly susceptible to these changes and promotes carcinogenesis. Future investigations will further throw light on this new cancer causing phenomenon.
Conclusion
After many years of continued progress in the molecular characterization of human cancers, a few marker models have been developed for clinical use. Microsatellite instability offers a good prognostic marker associated with different cancer types. The molecular detection of MSI is relatively simple in comparison to the identification of the majority of molecular genetic characteristics of potential clinical value, such as gene mutations and alterations in gene expression. Knowledge of the clinicopathological characteristics and other causative agents may facilitate the use of MSI detection as an integral part of the routine classification of all gastrointestinal tumours in the future. MSI based approach will provide a wealth of opportunities for analyzing the applicability of molecular characterization of cancer and exploring the possible benefits of its integration with other traditional approaches.
References
- Alterations of tumour suppressor and tumour-related genes in the development and progression of gastric cancer. World J Gastroenterol. 2006;12:192-8.
- [Google Scholar]
- The two histological main types of gastric carcinoma: Diffuse and so-called intestinal-type carcinoma.An attempt at a histo-clinical classification. Acta Pathol Microbiol Scand. 1965;64:31-49.
- [Google Scholar]
- Genetic classification of intestinal-type and diffuse-type gastric cancers based on chromosomal loss and microsatellite instability. Virchows Arch. 2003;443:491-500.
- [Google Scholar]
- Microsatellite variability differs between dinucleotide repeat motifs- evidence from Drosophila melanogaster. Mol Biol Evol. 2000;17:1277-85.
- [Google Scholar]
- The rise, fall and renaissance of microsatellites in eukaryotic genomes. Bioessays. 2006;28:1040-50.
- [Google Scholar]
- Mechanistic basis for microsatellite instability. In: Goldstein DB, Schlotterer C, eds. Microsatellites: Evolution and applications. Oxford: Oxford University Press; 1999. p. :34-48.
- [Google Scholar]
- Destabilization of tracts of simple repetitive DNA in yeast by mutations affecting DNA mismatch repair. Nature. 1993;365:274-6.
- [Google Scholar]
- Climatic effect on microsatellite diversity in wild emmer wheat, Triticum dicoccoides, at Yehudiyya microsite, Israel. Heredity. 2002;89:127-32.
- [Google Scholar]
- High-resolution fluorescent analysis of microsatellite instability in gastric cancer. Euro J Gastroenterol Hepatol. 2007;19:701-9.
- [Google Scholar]
- Genetic instability of microsatellites in endometerial carcinoma. Cancer Res. 1993;53:5100-3.
- [Google Scholar]
- p53 mutations and microsatellite instability in sporadic gastric cancer: when guardians fail. Cancer Res. 1994;54:4750-5.
- [Google Scholar]
- A National Cancer Institute workshop on microsatellite instability for cancer detection and familial predisposition: development of international criteria for the determination of microsatellite instability in colorectal cancer. Cancer Res. 1998;58:5248-57.
- [Google Scholar]
- Two modes of microsatellite instability in humancancer: differential connection of defective DNA mismatch repair to dinucleotide repeat instability. Nucleic Acids Res. 2005;33:1628-36.
- [Google Scholar]
- Microsatellite instability in gastric cancer is associated with better prognosis in only stage II cancers. Surgery. 2006;139:347-56.
- [Google Scholar]
- Marked differences in the frequency of microsatellite instability in gastric cancer from different countries. Am J Gastroenterol. 1999;94:3034-8.
- [Google Scholar]
- Microsatellite instability, prognosis and metastasis in gastric cancers from a low risk population. Int J Cancer. 2000;89:444-52.
- [Google Scholar]
- Microsatellite instability is a genetic marker for the development of multiple gastric cancers. Int J Cancer. 2001;95:350-3.
- [Google Scholar]
- Inactivation of the type II TGF-β receptor in colon cancer cells with microsatellite instability. Science. 1995;268:1336-8.
- [Google Scholar]
- Microsatellite instability in the insulin-like growth factor II receptor gene in gastrointestinal tumours. Nat Genet. 1996;14:255-7.
- [Google Scholar]
- Gastric cancer with high level microsatellite instability: target gene mutations, clinicopathologic features and long term survival. Hum Pathol. 2008;39:925-32.
- [Google Scholar]
- Microsatellite instability in synchronous gastric carcinomas. Int J Cancer. 2001;91:619-24.
- [Google Scholar]
- Mutational analysis of mononucleotide repeats in dual specificity tyrosine phosphatase genes in gastric and colon carcinomas with microsatellite instability. APMIS. 2010;118:389-93.
- [Google Scholar]
- High EPHB2 mutation rate in gastric but not endometrial tumours with microsatellite instability. Oncology. 2007;26:308-11.
- [Google Scholar]
- Frequent frameshift mutations of RIZ in sporadic gastrointestinal and endometrial carcinomas with microsatellite instability. Cancer Res. 2000;60:4701-4.
- [Google Scholar]
- MRE11 expression is impaired in gastric cancer with microsatellite instability. Carcinogenesis. 2004;25:2337-43.
- [Google Scholar]
- Somatic mutations in the DNA damage-response genes ATR and CHK1 in sporadic stomach tumours with microsatellite instability. Cancer Res. 2001;61:7727-30.
- [Google Scholar]
- Mutational and expressional analysis of RFC3, a clamp loader in DNA replication, in gastric and colorectal cancers. Hum Pathol. 2010;41:1431-7.
- [Google Scholar]
- Germline and somatic mutations in hMSH6 and hMSH3 in gastrointestinal cancers of the microsatellite mutator phenotype. Gene. 2001;272:301-13.
- [Google Scholar]
- Frameshift mutations of Wnt pathway genes AXIN2 and TCF7L2 in gastric carcinomas with high microsatellite instability. Hum Pathol. 2009;40:58-64.
- [Google Scholar]
- Microsatellite alterations and target gene mutations in the early stages of multiple gastric cancer. J Pathol. 2001;194:334-40.
- [Google Scholar]
- Frameshift mutations of autophagy-related genes ATG2B, ATG5, ATG9B and ATG12 in gastric and colorectal cancers with microsatellite instability. J Pathol. 2009;217:702-6.
- [Google Scholar]
- Somatic frameshift mutations in the Bloom syndrome BLM gene are frequent in sporadic gastric carcinomas with microsatellite mutator phenotype. BMC Genet. 2001;2:1471-7.
- [Google Scholar]
- Somatic frameshift mutations in the BAX gene in colon cancers of the microsatellite mutator phenotype. Science. 1997;275:967-9.
- [Google Scholar]
- Instabilotyping: comprehensive identification of frameshift mutations caused by coding region microsatellite instability. Cancer Res. 2001;61:6046-9.
- [Google Scholar]
- Mechanisms of inactivation of the receptor tyrosine kinase EPHB2 in colorectal tumours. Cancer Res. 2005;65:10170-3.
- [Google Scholar]
- Promoter methylation of TGFβ receptor I and mutation of TGF β receptor II are frequent events in MSI sporadic gastric carcinomas. J Pathol. 2003;200:32-8.
- [Google Scholar]
- The clinicopathological features of gastric carcinomas with microsatellite instability may be mediated by mutations of different target genes-a study of TGFβRII, IGFIIR and BAX genes. Am J Pathol. 1998;153:1211-9.
- [Google Scholar]
- Microsatellite instability and mutation analysis of candidate genes in urothelial cell carcinomas of upper urinary tract. Oncogene. 2006;25:2113-8.
- [Google Scholar]
- Hypersensitivity of tumour cell lines with microsatellite instability to DNA double strand break producing chemotherapeutic agent bleomycin. Cancer Res. 2004;64:4760-7.
- [Google Scholar]
- Altered expression of DNA-dependent protein kinase catalytic subunit (DNA-PKcs) during gastric carcinogenesis and its clinical implications on gastric cancer. Int J Oncol. 2007;31:859-66.
- [Google Scholar]
- Increased beta-catenin mRNA levels and mutational alterations of the APC and beta-catenin gene are present in intestinal-type gastric cancer. Carcinogenesis. 2002;23:87-91.
- [Google Scholar]
- EphB2 expression across 138 human tumour types in a tissue microarray: high levels of expression in gastrointestinal cancer. Clin Cancer Res. 2005;11:6450-8.
- [Google Scholar]
- Detection of frameshift mutations of RIZ in gastric cancers with microsatellite instability. World J Gastroenterol. 2004;10:2719-22.
- [Google Scholar]
- Microsatellites: simple sequences with complex evolution. Nat Rev Genet. 2004;5:435-45.
- [Google Scholar]
- Defective mismatch repair, microsatellite mutation bias, and variability in clinical cancer phenotypes. Cancer Res. 2010;70:431-5.
- [Google Scholar]
- The promise and perils of Wnt signaling through beta-catenin. Science. 2002;296:1644-6.
- [Google Scholar]
- Mutations in AXIN2 cause colorectal cancer with defective mismatch repair by activating betacatenin/TCF signalling. Nat Genet. 2000;26:146-7.
- [Google Scholar]
- Mutations within Wnt pathway genes in sporadic colorectal cancers and cell lines. Int J Cancer. 2006;119:1837-42.
- [Google Scholar]
- Genetic alterations in the human Tcf-4 gene in Japanese patients with sporadic gastrointestinal cancers with microsatellite instability. Oncology. 2001;61:156-61.
- [Google Scholar]
- The relationship between frameshift mutations of transforming growth factor-β type II receptor, insulin growth factor II receptor, bcl-2 associated X protein, E2F4 and microsatellite instability in gastric carcinoma. Zhongua Wai Ke Za Zhi. 2006;44:344-8.
- [Google Scholar]
- Hereditary multiple fibrofolliculomas with trichodiscomas and achrochordons. Arch Dermatol. 1977;113:1674-7.
- [Google Scholar]
- Mutations in a novel gene lead to kidney tumours, lung wall defects and benign tumours of the hair follicle in patients with Birt-Hogg-Dube syndrome. Cancer Cell. 2002;2:157-64.
- [Google Scholar]
- Birt-Hogg-Dubé (BHD) gene mutations in human gastric cancer with high frequency microsatellite instability. Cancer Lett. 2007;248:103-11.
- [Google Scholar]
- Frameshift mutation of UVRAG, an autophagy-related gene, in gastric carcinomas with microsatellite instability. Hum Pathol. 2008;39:1059-63.
- [Google Scholar]
- Concerted promoter hypermethylation of hMLH1, p16INK4A, and E-cadherinin gastric carcinomas with microsatellite instability. J Pathol. 2003;200:23-31.
- [Google Scholar]
- CDH1 gene promoter hypermethylation in gastric cancer. Diagn Mol Pathol. 2006;15:24-9.
- [Google Scholar]
- Microsatellite instability: the mutator that mutates the other mutator. Nature Med. 1996;2:630-1.
- [Google Scholar]
- Correlation of microsatellite instability at multiple loci with long term survival in advanced gastric carcinoma. Arch Surg. 2009;144:722-7.
- [Google Scholar]
- Microsatellite instability in gastric intestinal metaplasia in patients with and without gastric cancer. Am J Pathol. 2000;156:537-43.
- [Google Scholar]
- Clinicopathologic implications of genetic instability in intestinal-type gastric cancer and intestinal metaplasia as a precancerous lesion. Am J Clin Pathol. 2008;129:613-21.
- [Google Scholar]
- Clinical usefulness of microsatellite instability for the prediction of gastric adenoma or adenocarcinoma in patients with chronic gastritis. Br J Cancer. 2000;82:1814-8.
- [Google Scholar]
- Stage IV early gastric cancer: two cases with microsatellite instability. Langenbecks Arch Surg. 2008;393:105-9.
- [Google Scholar]
- Distinct clinical features and outcomes of gastric cancers with microsatellite instability. Mod Pathol. 2002;15:632-40.
- [Google Scholar]
- The role of histological investigation in prognostic evaluation of advanced gastric cancer. Analysis of histological structure and molecular changes compared with invasive pattern and stage. Virchows Arch. 2001;439:158-69.
- [Google Scholar]
- Differences in genomic instability between intestinal- and diffuse- type gastric cancer. Gastric Cancer. 2005;8:238-44.
- [Google Scholar]
- Assessment of microsatellite alterations in young patients with gastric adenocarcinoma. Cancer. 1997;79:684-7.
- [Google Scholar]
- Medullary-type poorly differentiated adenocarcinoma of the large bowel: a distinct clinicopathologic entity characterized by microsatellite instability and improved survival. J Clin Oncol. 1999;17:2429-38.
- [Google Scholar]
- Clinicopathological and molecular characteristics of gastric and colorectal carcinomas in elderly. Pathol Int. 2007;57:303-14.
- [Google Scholar]
- Age-related hypermethylation of the hMLH1 promoter in gastric cancers. Int J Cancer. 2001;94:208-11.
- [Google Scholar]
- Role of methylation of the hMLH1 gene promoter in the development of gastric and colorectal carcinoma in the elderly. Geriatr Gerontol Int. 2010;10:S207-12.
- [Google Scholar]
- Clinicopathologic and genetic characteristics of gastric cancer in young male and female patients. Oncol Rep. 2006;16:11-5.
- [Google Scholar]
- World Health Organization classification of tumours. Pathology and genetics of tumours of the digestive system. Lyon: IARC Press; 2000. p. :37-67.
- Clinicopathologic characteristics and surgical outcomes of mucinous gastric carcinoma. Ann Surg Oncol. 2006;13:836-42.
- [Google Scholar]
- Mucin phenotype and microsatellite instability in early multiple gastric cancers. Int J Cancer. 2002;100:419-24.
- [Google Scholar]
- Mutations at coding repeat sequences in mismatch repair-deficient human cancers: toward a new concept of target genes for instability. Cancer Res. 2002;62:2447-54.
- [Google Scholar]
- Pathogenesis of DNA repair-deficient cancers: a statistical meta-analysis of putative Real Common Target genes. Oncogene. 2003;22:2226-35.
- [Google Scholar]
- Red meat, family history, and increased risk of gastric cancer with microsatellite instability. Cancer Res. 2001;61:5415-9.
- [Google Scholar]
- Effects of dietary intake and genetic factors on hypermethylation of the hMLH1 gene promoter in gastric cancer. World J Gastroenterol. 2005;25:3834-41.
- [Google Scholar]
- Quantification of epigenetic and genetic 2nd hits in CDH1 during hereditary diffuse gastric cancer syndrome progression. Gastroenterology. 2009;136:2137-48.
- [Google Scholar]
- MSI phenotype and MMR alterations in familial and sporadic cancer. Int J Cancer. 2011;128:1606-13.
- [Google Scholar]
- Microsatellite instability and gastric non invasive neoplasia in a high risk population in Cesena, Italy. J Clin Pathol. 2005;58:805-10.
- [Google Scholar]
- Microsatellite instability in Japanese Vs European American patients with gastric cancer. Arch Surg. 2002;137:960-6.
- [Google Scholar]
- Chemosensitivity and survival in gastric cancer patients with microsatellite instability. Ann Surg Oncol. 2009;16:2510-5.
- [Google Scholar]
- Allelic imbalance at p53 and microsatellite instability are predictive markers for resistance to chemotherapy in gastric carcinoma. Ann Surg Oncol. 2009;16:2926-35.
- [Google Scholar]
- CpG island metylation status in gastric carcinoma with and without infection of Epstein Barr Virus. Clin Cancer Res. 2006;12:2995-3002.
- [Google Scholar]
- Epstein Barr Virus, p53 protein and microsatellite instability in the adenoma carcinoma sequence of the stomach. Hum Pathol. 2002;33:415-20.
- [Google Scholar]
- Gastric carcinoma with lymphoid stroma: its morphologic characteristic and prognostic correlations. Cancer. 1976;38:232-43.
- [Google Scholar]
- Intratumour T cells, their activation status and survival in gastric carcinomas characterised for microsatellite instability and Epstein Barr Virus infection. Virchows Arch. 2006;448:344-53.
- [Google Scholar]
- Lymphocyte rich gastric cancer: associations with Epstein Barr Virus, microsatellite instability, histology and survival. Mod Pathol. 2003;16:641-51.
- [Google Scholar]
- Helicobacter pylori and gastrointestinal tract adenocarcinoma. Nat Rev Cancer. 2002;2:28-37.
- [Google Scholar]
- Helicobacter pylori impairs DNA mismatch repair in gastric epithelial cells. Gastroenterology. 2002;123:542-53.
- [Google Scholar]
- Helicobacter pylori and EBV in gastric carcinomas: methylation status and microsatellite instability. World J Gastroenterol. 2010;21:312-9.
- [Google Scholar]
- Demonstration and characterization of mutations induced by Helicobacter pylori organisms in gastric epithelial cells. Helicobacter. 2006;11:272-86.
- [Google Scholar]
- Helicobacter pylori infection induces genetic instability of nuclear and mitochondrial DNA in Gastric cells. Clin Cancer Res. 2009;15:2995-3002.
- [Google Scholar]
- Helicobacter pylori infection generates genetic instability in gastric cells. Biochim Biophys Acta. 2010;1806:58-65.
- [Google Scholar]
- Analysis of the differences in structural chromosomal aberrations of the gastric mucosa between H. pylori positive and negative gastric cancer patients: involvement of H. pylori in the onset of gastric cancer and examination of the mechanism in gastric carcinogenesis following H. pylori eradication. Oncol Rep. 2006;16:1333-42.
- [Google Scholar]
- Mitochondrial D-loop 3’ (CA)n repeat polymorphism: optimization of analysis and population data. Electrophoresis. 1997;18:2857-60.
- [Google Scholar]
- Length heteroplasmy in the first hypervariable segment of the human mtDNA control region. Am J Hum Genet. 1995;57:248-56.
- [Google Scholar]
- Mitochondrial gene mutation, but not large-scale deletion, is a feature of colorectal carcinomas with mitochondrial microsatellite instability. Int J Cancer. 1999;83:625-9.
- [Google Scholar]
- Mutation in D-Loop region of mitochondrial DNA in gastric cancer and its significance. World J Gastroenterol. 2005;11:3304-6.
- [Google Scholar]
- Helicobacter pylori-induced oxidative stress and DNA damage in a primary culture of human gastric mucosal cells. Dig Dis Sci. 2002;47:1405-12.
- [Google Scholar]
- Microsatellite instability, mitochondrial DNA large deletions, and mitochondrial DNA mutations in gastric carcinoma. Genes Chromosomes Cancer. 2001;32:136-43.
- [Google Scholar]
- Tumoral cell mtDNA ~8.9 kb deletion is more common than other deletions in gastric cancer. Arch Med Res. 2006;37:848-53.
- [Google Scholar]
- Mitochondrial microsatellite instability in gastric cancer and gastric epithelial dysplasia as a precancerous lesion. Cancer Epidemiol. 2010;34:323-7.
- [Google Scholar]
- Association between Helicobacter pylori-related peptic ulcer tissue and somatic mitochondrial DNA mutations. Clin Chem. 2007;53:1390-2.
- [Google Scholar]
- Phylogenesis and regulated expression of the RUNT domain transcription factors RUNX1 and RUNX3. Blood Cells Mol Dis. 2003;30:161-3.
- [Google Scholar]
- Promoter methylation status of DAP-kinase and RUNX3 genes in neoplastic and non-neoplastic gastric epithelia. Cancer Sci. 2003;94:360-4.
- [Google Scholar]
- Frequent loss of RUNX3 expression by promoter hypermethylation in gastric carcinoma. Pathobiology. 2004;71:137-43.
- [Google Scholar]
- Prognostic value and clinicopathologicalprofile of microsatellite instability in gastric cancer. Clin Cancer Res. 1998;4:1749-54.
- [Google Scholar]
- , Microsatellite instability in gastrointestinal tract tumors. Int J Surg Investig. 2000;2:267-74.
- [Google Scholar]
- Aberrant methylation within RUNX3 CpG island associated with the nuclear and mitochondrial microsatellite instability in sporadic gastric cancers. Results of a GOIM (Gruppo Oncologico dell’Italia Meridionale) prospective study. Ann Oncol. 2007;18:vi103-9.
- [Google Scholar]




