
Translate this page into:
Constitutive expression of SMAR1 confers susceptibility to Mycobacterium tuberculosis infection in a transgenic mouse model
Reprint requests: Dr Samit Chattopadhyay, National Centre for Cell Science (NCCS), NCCS complex, University of Pune Campus, Ganeshkhind, Pune 411 007, Maharashtra, India e-mail: samit@nccs.res.in
Dr Umesh D. Gupta, National JALMA Instiute for Leprosy & Other Mycobacterial Diseases (ICMR), Dr. M. Miyazaki Marg, PO Box 101, Tajganj, Agra 282 001, Uttar Pradesh, India e-mail: gupta.umesh95@gmail.com
-
Received: ,
This is an open access article distributed under the terms of the Creative Commons Attribution NonCommercial ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non commercially, as long as the author is credited and the new creations are licensed under the identical terms.
This article was originally published by Medknow Publications & Media Pvt Ltd and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Background & objectives:
Studies involving animal models of experimental tuberculosis have elucidated the predominant role of cytokines secreted by T cells and macrophages to be an essential component of the immune response against Mycobacterium tuberculosis infection. The immune activities of CD4+ T cells are mediated in part by Th1 cytokine interferon gamma (IFN-γ) which is produced primarily by T cells and natural killer (NK) cells and critical for initiating the immune response against intracellular pathogen such as M. tuberculosis. Nuclear matrix protein SMAR1 plays an important role in V(D)J recombination, T helper cell differentiation and inflammatory diseases. In this study a transgenic mouse model was used to study the role of SMAR1 in M. tuberculosis infection.
Methods:
Wild type BALB/c, C57BL/6, BALB/c-EGFP-SMAR1 and C57BL/6-SMAR1 transgenic mice were infected with M. tuberculosis (H37Rv). A dose of 100 bacilli was used for infection via respiratory route. Bacterial load in lung and spleen of infected mice was determined at 2, 4, 6 and 8 wk post-infection. Gene expression analysis for Th1 cytokines and inducible nitric oxide synthase (iNOS) was performed in infected lung tissues by quantitative reverse transcription (RT)-PCR.
Results:
SMAR1 transgenic mice from both BALB/c and C57BL/6 genetic background displayed higher bacillary load and susceptibility to M. tuberculosis infection compared to wild type mice. This susceptibility was attributed due to compromised of Th1 response exhibited by transgenic mice.
Interpretation & conclusions:
SMAR1 transgenic mice exhibited susceptibility to M. tuberculosis infection in vivo irrespective of genetic background. This susceptibility was attributed to downregulation of Th1 response and its hallmark cytokine IFN-γ. Hence, SMAR1 plays an important role in modulating host immune response after M. tuberculosis infection.
Keywords
Cytokine
IFN-γ transgenic
SMAR1
tuberculosis

Tuberculosis is one of the major causes of morbidity and mortality worldwide, with approximately one-third of the world's population exposed to it1. The prevalence and persistence of Mycobacterium tuberculosis infection manifest a unique model of pathogen-host coexistence. The pathogenesis of tuberculosis relies upon the dynamic interplay between bacterial virulence and host resistance2. Studies on animal models and gene-disrupted mice have shown that the protective immunity against M. tuberculosis depends on a wide range of innate and adaptive immune mechanisms2345. Studies using mouse model have demonstrated a key role of T type 1 (Th1) CD4+ cells with the aid of CD8+ cells678. This can be evidenced by the studies demonstrating that mice deficient in CD4 and major histocompatibility complex-II (MHC-II) exhibit increased mycobacterial burden and decreased survival after infection46. Following exposure, alveolar macrophages engulf the bacilli leading to a localized pulmonary immune response, followed by alveolitis of activated α/β T-cell receptor-positive lymphocytes and macrophages recruitment. This sequence of events leads to establishment of a strong M. tuberculosis antigen-specific Th1 response, with large amounts of locally secreted interferon gamma (IFN-γ) to control bacterial growth67. The ability of CD4+T cells to produce IFN-γ, which activates macrophages to contain the intracellular pathogen, is central to this protection. Interferon-γ activates macrophages and promotes intracellular bacterial killing by permitting phagosomal maturation and production of antimicrobial reactive nitrogen intermediates and reactive oxygen intermediates4910. The importance of IFN-γ signaling axis is evident from the fact that the mice lacking IFN-γ, IFN-γ receptor or signal transducer and activator of transcription 1 (STAT1) are extremely susceptible to M. tuberculosis infection111213. Another Th1 cytokine, interleukin-12 (IL-12) has been shown to promote T cell differentiation towards Th1 also plays an important role. The secretion of IL-12 has immunopotentiation effects in terms of its ability to initiate the development of Th1 phenotype in naive T cells and the ability to augment IFN-γ production in antigen-activated Th1 cells1415. This is evident by the fact that individuals with deficiencies in IFN-γ and IL-12 signaling pathways exhibit increased risk of tuberculosis13. IL-12 is known to induce CD4+ and CD8+ T cells to secrete IFN-γ via STAT4 dependent pathways. However, IFN-γ can be produced by a variety of immune cells including MHC-II restricted CD4+ T cells, MHC-I restricted CD8+T cells, CD1 restricted natural killer (NK)T cells and certain types of dendritic cells or macrophages61415. In addition, the importance of host nuclear proteins and transcription factors has also been reported. The Th1 specific transcription factor T-bet deficiency leads to susceptibility to M. tuberculosis infection in mice16, nuclear protein BAT3 is shown to be associated with the fate of antigen presenting cells during infection17, and apoptotic protein Bim has been shown to be the master regulator of apoptosis induced by M. tuberculosis in macrophages18. Similarly, AIM2 (absent in melanoma-2) is responsible for the activation of inflammasome and host immune responses against M. tuberculosis. AIM2-deficient mice are highly susceptible to intratracheal infection with M. tuberculosis and this is associated with defective IL-1β and IL-18 production together with impaired Th1 responses19.
We have previously identified SMAR1 (scaffold/matrix attachment region binding protein 1) from mouse double positive thymocytes screen as nuclear matrix associated protein that interacts with cis regulatory matrix attachment regions (MARs) present in the genome20. SMAR1 functions as negative regulator of T cell receptor beta (TCRβ) locus, it represses transcription from Eβ enhancer and regulates V(D)J recombination process which is critical for T cell diversity and response2122. Additionally, SMAR1 is involved in a number of cellular processes and transcriptional regulation of several genes23. Subsequent studies on transgenic mice established the critical role of SMAR1 in T cell development and maturation. As shown previously, the constitutive expression of SMAR1 in mice leads to perturbed development of lymphoid organs with pronounced splenomegaly and lymphadenopathy accompanied by follicular hyperplasia, indicating a hyper-responsive immune system22. Recently, the role of SMAR1 in inflammatory diseases like asthma and colitis has been established2425. We have earlier shown that SMAR1 inhibits Th1 differentiation of naïve CD4+ T cells and SMAR1 transgenic mice demonstrate impaired Th1 and Th17 responses2425. Given the phenotype of transgenic mice observed in our previous studies, in this study we utilized these mice for M. tuberculosis infection to evaluate the role of SMAR1 in a disease where Th1 response is primary for the containment of infection.
Material & Methods
All animal experiments were conducted in the BSL-III laboratory at the National JALMA Institute for Leprosy and Other Mycobacterial Diseases (NJIL&OMD), Agra, India, as per protocols and guidelines approved by the Institutional Animal Ethics Committees of NJIL&OMD, Agra, and National Center for Cell Science (NCCS), Pune, India, during December 2009 and November 2013.
Mice and genotyping: Wild type BALB/c, BALB/c-EGFP-SMAR1, C57BL/6, and C57BL/6-SMAR1 transgenic mice were bred and maintained under specific pathogen free conditions at animal facility of National Institute of Immunology, New Delhi, and NCCS, Pune, India. Homozygous BALB/c-EGFP-SMAR1 transgenic and C57BL/6-SMAR1 transgenic lines were generated at National Institute of Immunology, New Delhi, as described previously2226. For genotyping of SMAR1 transgenic mice, the tail DNA from litters was extracted. Briefly, a 2-mm tail biopsy was incubated in high salt digestion buffer containing 50 mM Tris-HCl, 100 mM ethylene dimaine tetra acetic acid (EDTA), 0.5 per cent sodium dodecyl sulphate (SDS), and 0.5 mg/ml proteinase K for 16 h at 55 °C. The lysates were processed for DNA isolation by phenol/chloroform extraction method followed by ethanol precipitation. DNA was subjected to PCR analysis using primers (Table) spanning CMV promoter and gene coding junction. The positively scored mice were also analyzed for the presence of transgene by Southern hybridization using CMV promoter specific probes as described previously22.
Mycobacterial strain: The virulent M. tuberculosis strain (H 37 R v) was obtained from Mycobacterial Repository at NJIL&OMD, Agra. Mycobacteria were cultured in Middlebrook 7H9 broth (BD Biosciences, USA) containing 0.5 per cent Tween-80 (Sigma-Aldrich, USA) supplemented with 10 per cent oleic acid albumin dextrose and catalase (OADC, BD Biosciences), until mid log phase and then the bacterial culture was stored in aliquots at -80° C, until further use. The turbidity of the cultures was adjusted to McFarland standard as described27 and the inoculum size was confirmed by titration and plating serial dilutions on Middlebrook 7H11 plates supplemented with 10 per cent OADC enrichment. The plates were incubated at 37°C for 4 wk prior to counting of colonies.
Aerosol infections: Both wild type and SMAR1 transgenic mice (age 6-8 wk) of either sex were infected with M. tuberculosis via respiratory route using a closed chamber inhalation exposure system (Glas-Col, USA). For each time point four mice per group were taken. Mice were exposed for 45 min cycle to nebulize bacteria at pre-calibrated density, optimized to deliver a standard low dose of approximately 100 bacilli. For all infections, the actual infection dose was confirmed by plating total lung homogenate from a minimum of two mice on Middlebrook 7H11 nutrient medium after 24 h of aerosol exposure.
Determination of bacterial burden: Bacterial burden in lung and spleen of wild type and SMAR1 transgenic mice was determined at different time points after aerosol infection. For this, lung and spleen were aseptically removed from euthanized animals of each group. Organs were homogenized in phosphate buffered saline (PBS) containing 0.05 per cent Tween-80 (pH 7.4). Serial dilutions of homogenized organs were plated in duplicates on Middlebrook 7H11 agar plates supplemented with 10 per cent OADC. Plates were incubated at 37°C for 21-28 days, colonies on plates were enumerated and results were expressed as log10 cfu per organ.
Enumeration of bacilli: Lung tissues from each experimental group of euthanized mice were fixed with 10 per cent neutral buffered formalin. To visualize the acid-fast bacilli (AFB), tissue sections were stained using Ziehl-Neelsen method and observed microscopically to count the acid-fast bacilli.
Semi-quantitative and quantitative reverse transcription (RT)-PCR: The lung lobes from infected mice were homogenized in one ml of TRIzol reagent (Invitrogen, USA). Samples were stored until use at -80°C, following lysis in TRIzol. Total RNA was isolated as per manufacturer's protocol by TRIzol/chloroform method. Semi-quantitative RT-PCR analysis was performed as described22, briefly the total RNA was isolated from cells and treated with RNase free DNase I (Roche, USA), to eliminate genomic DNA contamination. For each assay, 10 μg of total RNA was subjected to reverse transcription in a 20 μl reaction mixture containing 1X random hexanucleotide mix, 1 mM deoxynucleoside triphosphates, RNase inhibitor, and MuMLV reverse transcriptase (Invitrogen, USA). PCR was carried out in thermal cycler (Eppendorf, USA), in a 25 μl reaction mixture using 1:5 diluted 1 μl cDNA as template. RT-PCR products were separated on a 1.2 per cent agarose gel and visualized by staining with ethidium bromide. Band intensities were quantified using Image J software (NIH, USA) and normalized with respect to the actin loading control. Quantitative real time PCR performed as described24 in Master Cycler gradient (Eppendorf, Germany) using SYBR Green master mix (Eppendorf). The relative abundance of mRNA was determined by ΔΔCt, where ΔCt = Ct (gene) - Ct (GAPDH) and ΔΔCt = ΔCt (sample) – ΔCt (Calibrator). Total RNA from uninfected mice served as calibrator. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a normalizing control. The relative expressions of IFN-γ, IL-12 and iNOS were calculated as folds change over uninfected mice. The sequences of respective gene specific primers used are given in the Table.
ELISA: Total T cells were isolated from spleen using nylon wool column. CD4+ T cell were enriched using magnetically labelled CD4+ (L3T4) micro beads (Milteny Biotech, USA). Cells were cultured in RPMI (Roswell Park Memorial Institute) medium supplemented with 10 per cent foetal bovine serum (FBS) and stimulated by 1 μg/ml of anti-CD3 and 3 μg/ml of anti-CD28 antibodies (BD Biosciences) for 24 h. Culture supernatants were harvested and subjected to quantification of IFN-γ secretion using an ELISA kit (BD Biosciences), as per manufacturers instructions.
Statistical analysis: Statistical analysis between experimental groups was done using two-tailed Student t-test. The values are represented as mean ± standard error of mean (SEM) or standard deviation (SD).
Results
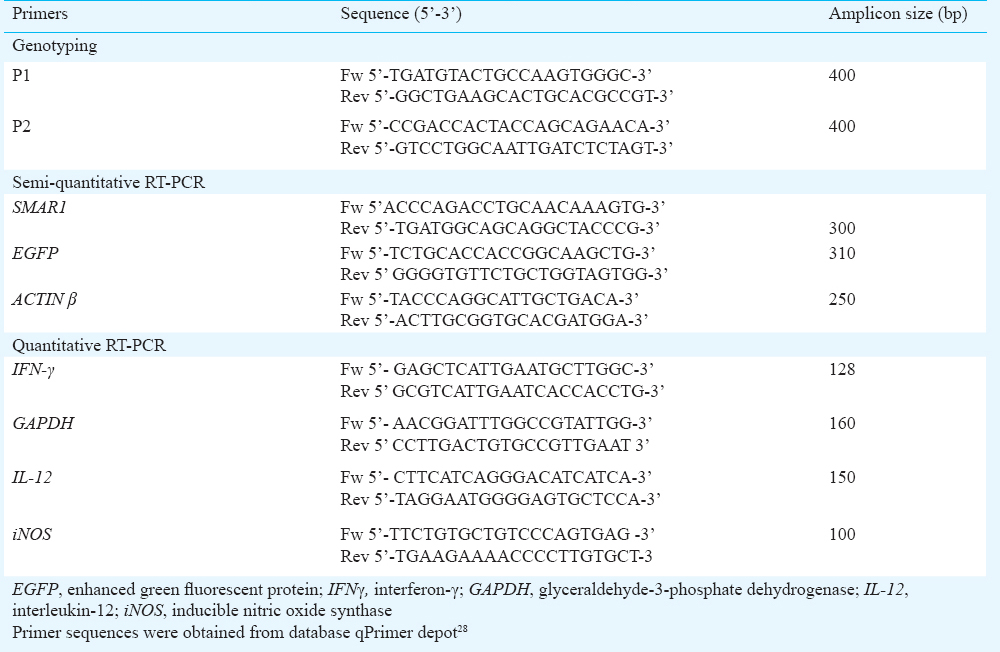
SMAR1 transgenic mice demonstrate perturbed immune response and downregulation of Th1 cytokine IFN-γ: Transgenic mice litters were genotyped for the presence of SMAR1 transgene by PCR on tail tissue genomic DNA. Fig. 1A demonstrates the genotype pattern of SMAR1 transgenic litters from both BALB/c and C57BL/6 genetic background. The presence of 400 bp amplicon using primers flanking CMV-promoter and gene coding junction, scored positive for the presence of transgene in a set of genotyped mice. We investigated the functional impact of SMAR1 overexpression by analyzing the T cell responses in transgenic mice litters. To confirm the overexpression of SMAR1 in transgenic mice, we evaluated the mRNA level of SMAR1 in different tissues of wild type and transgenic mice. Semi-quantitative RT-PCR shown in Fig. 1B and C, demonstrated the overexpression of SMAR1 transcript in different tissues such as lymph node, thymus and spleen from BALB/c-EGFP-SMAR1 and C57BL/6-SMAR1 transgenic mice. The densitometry analysis of RT-PCR products indicated that SMAR1 expression was 4-5 fold higher in transgenic mice compared to the wild type counterparts (Fig. 1D and E). Further, ex vivo analysis of splenic CD4+ T cells from wild type and SMAR1 transgenic mice was performed. As demonstrated by quantitative RT-PCR (Fig. 1F), a significant reduction of ~3-4 folds in IFN-γ transcript was observed in anti-CD3/anti-CD28 stimulated CD4+ T cells from transgenic mice compared to wild type activated cells. Further quantification of secreted IFN-γ was performed in culture supernatants of anti-CD3/anti-CD28 stimulated CD4+ T cells from SMAR1 transgenic and wild type mice (Fig. 1G). The ELISA (Fig. 1G), revealed that SMAR1 transgenic CD4+ T cells produced substantially less IFN-γ (200-300 pg/ml) compared to wild type CD4+ T cells (~600-750 pg/ml). These results suggested that SMAR1 transgenic mice though exhibited hyper-cellularity of T cells in lymphoid organs (as reported previously22), CD4+T cells were unable to initiate IFN-γ production in presence of activating stimuli. The down-regulation of IFN-γ expression in T cells suggested the need to further investigate the immune response in these mice to infection.
- Genotyping and expression analysis in SMAR1 transgenic mice (A) Genotyping of SMAR1 transgenic mice performed on tail tissue DNA by PCR, using primers from CMV promoter and gene coding junction. P1 and P2 denote the position of primers; presence 400 bp amplicon confirmed the presence of transgene in positively scored mice. Left panel, BALB/c-SMAR1 transgenic genotype patterns (Lane 1 pEGFP-SMAR1 plasmid template as positive control, lanes 2, 3, 5, 6 represent transgenic litters, lane 4 represents wild type mice and M denotes 1 kb DNA ladder). Right panel represent genotypes from C57BL/6-SMAR1 transgenic litters (lanes 1, 3, 4, 6 represent transgenic litters while lanes 2, 5 represent wild type litters). (B) SMAR1 expression in lymph node (LN), thymus (Thy), spleen (SP) from wild type BALB/c (lanes 1, 2, 3) and BALB/c-EGFP-SMAR1 transgenic (lanes 4, 5, 6). (C) SMAR1 expression in thymus (Thy) and spleen (SP) of wild type C57BL/6 (lanes 1, 2) and C57BL/6-SMAR1 transgenic mice (lanes 3, 4) by semi-quantitative RT-PCR. Actin β is monitored as a loading control. (D) & (E) Densitometry analysis of Fig. 1B & 1C, by Image J Software demonstrating the fold change in expression after normalization with actin loading control. Error bar represents ± SD from three indpendent experiments. (F) Expression analysis of IFN-γ mRNA by quantitative RT-PCR in splenic CD4+ T cells from wild type and SMAR1 transgenic mice stimulated with anti-CD3 and anti-CD28 ligands for 24 h. (G) ELISA demonstrating the secretion of IFN-γ by CD4+ T cells stimulated ex vivo with anti-CD3 and anti-CD28 ligands. Error bars represent ± SEM from three independent experiments (**P<0.01).
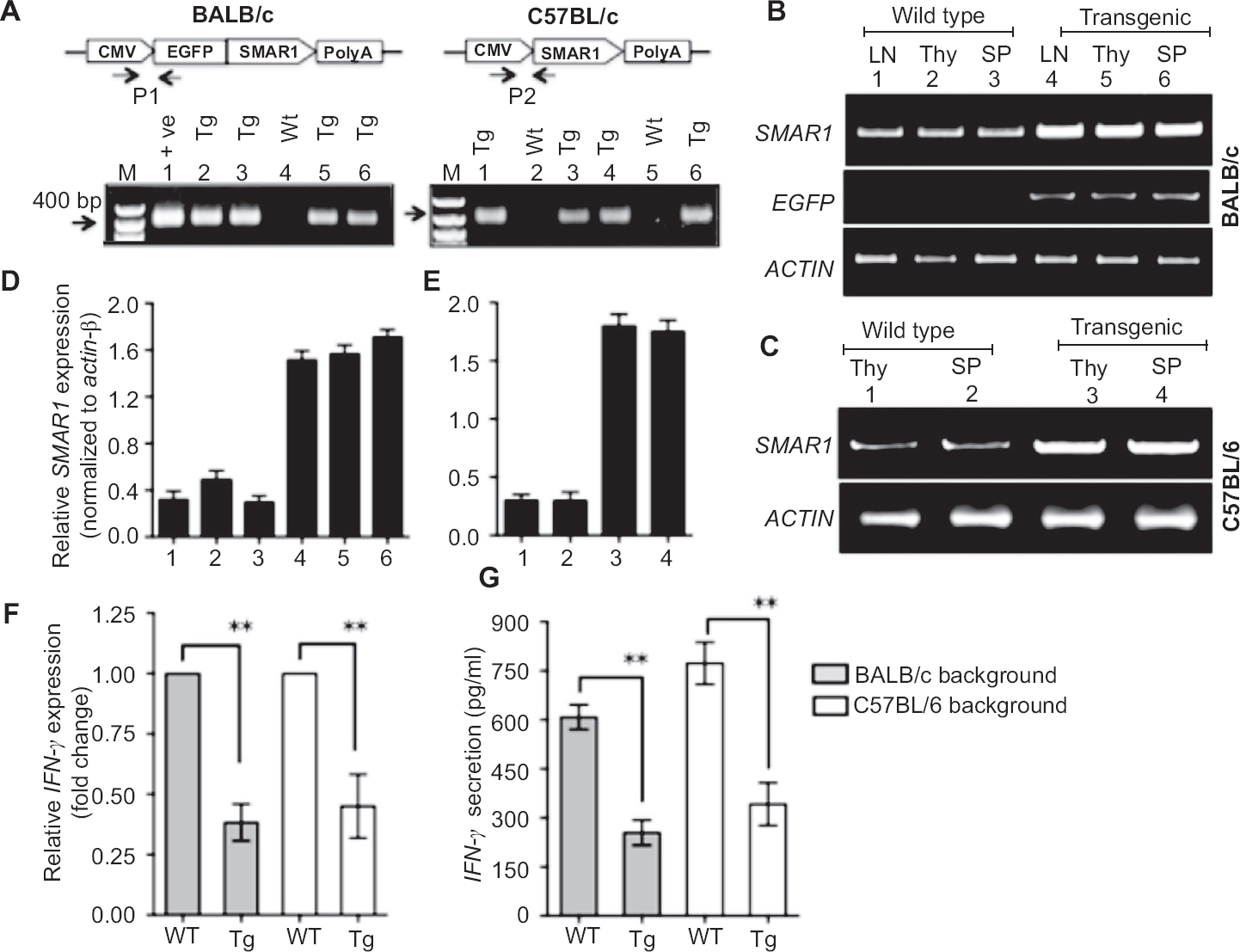
SMAR1 transgenic mice exhibit susceptibility to M. tuberculosis infection: To access the role of SMAR1 in protective immune response against M. tuberculosis, both wild type and SMAR1 transgenic mice of BALB/c and C57BL/6 genetic background were infected with low dose of virulent M. tuberculosis strain H37Rv via aerosol route. Following infection, mice were sacrificed after 2, 4, 6, 8 wk respectively to monitor the progression of infection. The bacterial burden in the target organs was determined, for which lung and spleen were aseptically removed, homogenized and the serial dilutions were plated on Middlebrook 7H11 agar plates. To ensure that the equal numbers of bacilli deposited in mice lung by aerosol challenge, each set of mice were analyzed next day after infection (taken as day zero) by plating their lung homogenates on 7H11 agar plates. The plates were incubated for 3-4 wk until the bacterial colonies appeared, the number of colonies on plates were counted for each sets of mice and expressed as log10 colony forming units (cfu). The day zero colony counts indicated that approximately 100-125 bacilli got implanted in the lungs of each set of mice after aerosol exposure. Further, the comparative analysis of colony forming units in lung and spleen was performed at different time points. After two weeks post infection, a substantial difference in bacterial burden was apparent in SMAR1 transgenic mice lungs compared to wild type mice lungs. After four weeks post infection, SMAR1 transgenic mice lungs exhibited ~1.6 to 1.8 fold more cfu than the lungs of wild type mice, which progressively increased up to six weeks (Fig. 2A and B). At eight weeks, there was a decline in bacterial numbers in wild type mice indicating a control of infection and induction of immune response. However, in SMAR1 transgenic mice the relative decrease in cfu at eight weeks was not significant as observed in case of wild type mice.
- Determination of bacterial burden in target organs of wild type and SMAR1 transgenic mice of BALB/c and C57BL/6 genetic backgrounds. Mice were infected with M. tuberculosis strain, H37Rv through aerosol. Mycobacterial burden was determined in lungs after 0, 2, 4, 6, 8 wk (A-B) and spleen (C-D) after 0, 4, 6 and 8 wk post infection. Data represent mean ± SD with four mice per group at each time point (*P<0.05).
In case of spleen, we analyzed bacterial burden from four weeks post infection as it has been previously shown that dissemination of bacilli from lung to other organs occurs in 2-3 wk time4. The SMAR1 transgenic mice spleen exhibited higher bacterial burden than wild type mice at all time points analyzed (Fig. 2C and D). The overall results from in vivo infection experiments and cfu analysis suggested that SMAR1 transgenic mice irrespective of genetic background exhibited more susceptibility to M. tuberculosis than the wild type mice
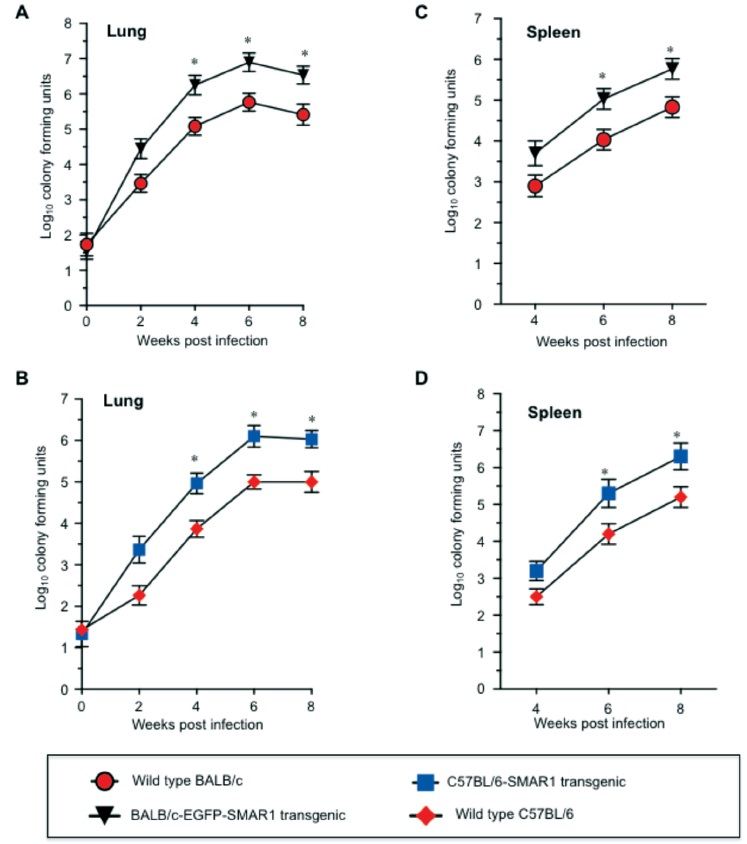
Histological analysis of infected tissues from wild type and SMAR1 transgenic mice: Phenotypic examination of lung and spleen at each time of sacrifice indicated distinct pathological differences between wild type and SMAR1 transgenic mice. The granulomatous lesions appeared moderately more visible with significant lymphocytic infiltrations in SMAR1 transgenic lung compared to wild type (data not shown). We then determined the bacilli count in the formalin fixed lung tissue sections by Ziehl-Neelsen staining. Slides were microscopically analyzed and numbers of acid-fast bacilli in tissue sections were enumerated. The analysis revealed higher AFB count in SMAR1 transgenic compared to wild type mice (Fig. 3).
- Plot demonstrating acid-fast bacilli (AFB) count in SMAR1 transgenic and wild type mice lungs. For quantification of bacilli in tissue sections 20 fields/slide were analyzed and average mean count was determined. Error bar represents average mean of five tissue sections from each animal (*P<0.05).
Susceptibility to infection is due to reduced IFN-γ expression and suppressed Th1 response in SMAR1 transgenic mice: We performed cytokine analysis in infected lungs from SMAR1 transgenic and wild type mice to determine the expression profile of genes, which are known to be critical determinants of Th1 response. Total mRNA was isolated from infected lung tissues subjected to quantitative RT-PCR and relative expression of IFN-γ, IL-12 and inducible nitric oxide synthase (i-NOS) transcripts were determined. In wild type mice an induction of IFN-γ expression was observed during the course of infection. However, in case of SMAR1 transgenic mice, IFN-γ transcript expressed at significantly (P<0.05) low levels compared to wild type mice (Fig. 4A). Additionally, we also observed a reduced expression of IL-12 and i-NOS in infected lungs of SMAR1 transgenic mice compared to wild type mice (Fig. 4B and C). These expression analyses suggested that transgenic mice, due to failure to mount effective Th1 response and inadequate IFN-γ expression, exhibited susceptibility to infection and higher bacillary load.
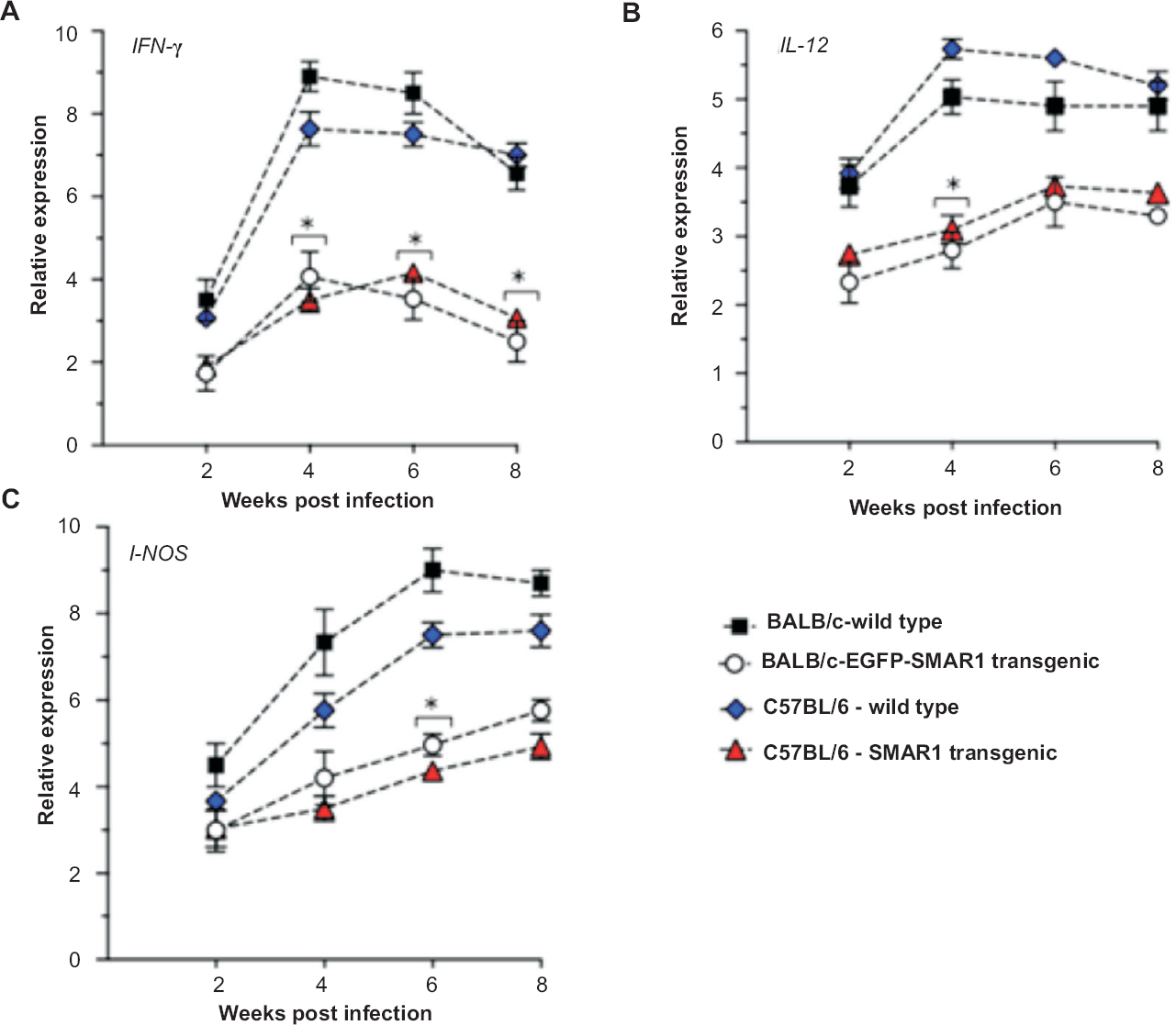
- Expression analysis of Th1 pathway genes in infected lung tissues. (A-C) Relative expression of IFN-γ, IL-12 and iNOS in infected lung samples from wild type and SMAR1 transgenic mice during the course of infection, determined by quantitative RT-PCR. The Y-axis denotes the relative expression represented as fold change after normalization with the values obtained from uninfected mice. GAPDH was used as an internal control. Values are represented as mean ± SD (n=4); *P<0.05.
Discussion
Tuberculosis is a complex disease and the success of M. tuberculosis as an intracellular pathogen is the outcome of its close association with the mammalian host. The studies on host-pathogen interactions have led to elucidate the multitude of signaling pathways and molecular event accounting for the cross-talk between the host and pathogen. There is a sensitive balance between the protective immunity and destructive pathology associated with the disease29. In a mouse model, the Th1 immunity generated in immunocompetent wild type mouse provides capacity to gain control of bacterial growth and holds infection at stationary level. Persistent stationary lung infection however, causes progressive lung pathology and eventually death. The failure of immunity to resolve the infection is consequence of generation of an inadequate level of Th1 immune response. Our study elucidates on the role of nuclear matrix protein SMAR1 in tuberculosis infection. We demonstrate that SMAR1 overexpression led to increased susceptibility to M. tuberculosis infection in a transgenic mouse model. Given that C57BL/6 mice are comparatively resistant to infection than BALB/c, the in vivo infection data convincingly demonstrated that irrespective of genetic background SMAR1 transgenic mice with C57BL/6 background displayed susceptibility to infection compared to wild type C57BL/6. SMAR1 transgenic mice exhibited substantially higher bacillary load in lungs and spleen than the wild type mice. It has been shown that aerosol infection results in an approximate 3-wk period of log linear growth in the lungs before further growth is inhibited and infection is stabilized by acquired immunity29. The long delay before the induction of Th1 adaptive immunity infection enables the infection to reach a stationary level that induces progressive and eventually lethal lung pathology3031. We also observed a similar trend in our infection studies where a progressive increase in cfu in lungs of both wild type and SMAR1 transgenic mice was observed up to six weeks and subsequent decrease in counts at eight weeks time, though the fold decline in cfu at late time points was comparatively less in SMAR1 transgenic lungs when compared to wild type mice. As reported in several studies before that immunity against M. tuberculosis is mediated by Th1 cells via the secretion of IFN-γ, but is elicited by macrophages through iNOS-dependent generation of nitric oxide and reactive nitrogen intermediates6910. The generation of Th1 mediated immunity however, is also dependent upon IL-12. Therefore, the magnitude of the Th1 response can be ascertained by determining changes in IL-12, IFN-γ and iNOS gene expression in the lungs during the course of infection910. It has been shown that IFN-γ knockout mice succumb to M. tuberculosis infection and mice die within four weeks of infection1112. Our results demonstrated that susceptibility of SMAR1 transgenic mice is attributed to their inefficiency to mount the protective Th1 response against the bacilli. Further, the CD4+ T cells from SMAR1 transgenic mice exhibited lower induction of IFN-γ in response to activating stimuli. Consistent with this, during the course of infection unlike wild type mice, no significant induction of IFN-γ was observed in the infected lung tissues of SMAR1 transgenic mice. This was similar to our earlier findings on SMAR1 transgenic mice showing that these mice had defective Th1 response as evidenced by downregulation of T-bet and lower IgG2a levels24. The Th1 specific transcription factor, T-bet is a positive regulator of IFN-γ transcription and T-bet knockout mice succumb to M. tuberculosis infection with high bacillary load and lower IFN-γ production16. In addition, data from our laboratory indicated the inability to SMAR1 transgenic CD4+ T cells to augment IFN-γ expression in response to mycobacterial antigens (unpublished data). Our microarray study also demonstrated the downregulation of chemokine and cytokine receptor genes in SMAR1 transgenic mice (unpublished data). This suggests that the perturbed Th1 response in SMAR1 transgenic mice may be in part due to the altered expression pattern of immune receptor genes, which is required for initiation of protective immune response to contain the infection. Given the basic function of SMAR1 in gene regulation, the comprehensive analyses of infected tissues at genome level are required. The whole genome studies such as RNA-sequencing, which utilize next generation sequencing coupled to epistasis analysis, can reveal more information on altered gene expression patterns and potential targets of SMAR1 during M. tuberculosis infection. This will also enable to dissect the specific signaling pathway that leads to perturb Th1 immune response and susceptibility of SMAR1 transgenic mice to infection. Which pathways are specific targets of SMAR1 remains to be studied using homozygous knockout mice. Studies in haplosufficiency state would help to delineate the specific targets and modes of regulation by SMAR1.
Overall this study shows that defective Th1 immune response in SMAR1 transgenic mice confers susceptibility to M. tuberculosis infection. Therefore, SMAR1 transgenic mice can be a model system to understand the fine tune control of Th1 response upon infections with intracellular pathogens.
Acknowledgment
Authors acknowledge the help and cooperation from Drs B. Ramanamurthy, Sijo V. Chemmannur, Surajit Sinha, Rahul M. Bankar and Shri Sunil Banskar. This study was funded by research grants from Department of Biotechnology (DBT), New Delhi. The first author (BY) is a recipient of fellowship from Lady TATA Memorial Trust, Mumbai, India. The second author (SKM) is a recipient of senior research fellowship from Indian Council of Medical Research (ICMR), New Delhi, India.
Conflicts of Interest: None.
References
- The balance between protective and pathogenic immune responses in the TB-infected lung. Nat Immunol. 2014;16:57-63.
- [Google Scholar]
- Innate and adaptive immune responses to human Mycobacterium tuberculosis infection. Tuberculosis (Edinb). 2009;89(Suppl 1):S77-80.
- [Google Scholar]
- Animal models of M. tuberculosis infection. Curr Protoc Microbiol 2007 10A.5.1-5.29. doi:10.1002/9780471729259.mc10a05s7
- [Google Scholar]
- Initiation and regulation of T-cell responses in tuberculosis. Mucosal Immunol. 2011;4:288-93.
- [Google Scholar]
- Cutting edge: Direct recognition of infected cells by CD4 T cells is required for control of intracellular Mycobacterium tuberculosis in vivo. J Immunol. 2013;191:1016-20.
- [Google Scholar]
- T lymphocytic and immature macrophage alveolitis in active pulmonary tuberculosis. J Infect Dis. 1996;173:1267-72.
- [Google Scholar]
- Role of innate cytokines in mycobacterial infection. Mucosal Immunol. 2011;4:252-60.
- [Google Scholar]
- Properties and protective value of the secondary versus primary T helper type 1 response to airborne Mycobacterium tuberculosis infection in mice. J Exp Med. 2005;201:1915-24.
- [Google Scholar]
- Disseminated tuberculosis in interferon gamma gene-disrupted mice. J Exp Med. 1993;178:2243-7.
- [Google Scholar]
- An essential role for interferon gamma in resistance to Mycobacterium tuberculosis infection. J Exp Med. 1993;178:2249-54.
- [Google Scholar]
- Inborn errors of IL-12/23- and IFN-gamma-mediated immunity: molecular, cellular, and clinical features. Semin Immunol. 2006;18:347-61.
- [Google Scholar]
- Interleukin-12 and tuberculosis: an old story revisited. Curr Opin Immunol. 2007;19:441-7.
- [Google Scholar]
- Compartmentalized bronchoalveolar IFN-gamma and IL-12 response in human pulmonary tuberculosis. Tuberculosis (Edinb). 2009;89:38-47.
- [Google Scholar]
- Increased susceptibility of mice lacking T-bet to infection with Mycobacterium tuberculosis correlates with increased IL-10 and decreased IFN-gamma production. J Immunol. 2005;175:4593-602.
- [Google Scholar]
- BAT3 regulates Mycobacterium tuberculosis protein ESAT-6-mediated apoptosis of macrophages. PLoS One. 2012;7:e40836.
- [Google Scholar]
- Bim is a crucial regulator of apoptosis induced by Mycobacterium tuberculosis. Cell Death Dis. 2014;5:e1343.
- [Google Scholar]
- Critical role of AIM2 in Mycobacterium tuberculosis infection. Int Immunol. 2012;24:637-44.
- [Google Scholar]
- SMAR1, a novel, alternatively spliced gene product, binds the Scaffold/Matrix-associated region at the T cell receptor beta locus. Genomics. 2000;68:93-6.
- [Google Scholar]
- SMAR1 and Cux/CDP modulate chromatin and act as negative regulators of the TCRbeta enhancer (Ebeta) Nucleic Acids Res. 2004;32:4862-75.
- [Google Scholar]
- Abnormal V(D)J recombination of T cell receptor beta locus in SMAR1 transgenic mice. J Biol Chem. 2005;280:9450-9.
- [Google Scholar]
- Gene regulation by SMAR1: Role in cellular homeostasis and cancer. Biochim Biophys Acta. 2011;1815:1-12.
- [Google Scholar]
- Nuclear matrix binding protein SMAR1 regulates T cell differentiation and allergic airway disease. Mucosal Immunol. 2015;8:1201-11.
- [Google Scholar]
- Regulation of T cell lineage commitment by SMAR1 during inflammatory and autoimmune diseases. Indian J Med Res. 2015;142:405-13.
- [Google Scholar]
- Transgenesis via permanent integration of genes in repopulating spermatogonial cells in vivo . Nat Methods. 2008;5:601-3.
- [Google Scholar]
- Mycobacterium indicus pranii as stand-alone or adjunct immunotherapeutic in treatment of experimental animal tuberculosis. Indian J Med Res. 2011;134:696-703.
- [Google Scholar]
- qPrimerDepot: a primer database for quantitative real time PCT. Nucleic Acid Res. 2007;35:805-9.
- [Google Scholar]
- For better or for worse: the immune response against Mycobacterium tuberculosis balances pathology and protection. Immunol Rev. 2011;240:235-51.
- [Google Scholar]
- Profiling early lung immune responses in the mouse model of tuberculosis. PLoS One. 2011;6:e16161.
- [Google Scholar]
- Long-term control of Mycobacterium tuberculosis infection is mediated by dynamic immune responses. J Immunol. 2005;175:1107-17.
- [Google Scholar]




