
Translate this page into:
Preventive action of curcumin in experimental acute pancreatitis in mouse
Reprint requests Dr Ke-Li Tian, Department of Biochemistry & Molecular Biology, Shandong University School of Medicine, 44# Wenhua Xi Road, Jinan 250012, Shandong, PR China tiankeli@sdu.edu.cn
-
Received: ,
This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-Share Alike 3.0 Unported, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This article was originally published by Medknow Publications & Media Pvt Ltd and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Background & objectives:
Curcuma longa (turmeric) has a long history of use in Ayurvedic medicine as a treatment for inflammatory conditions. The purpose of the present study was to investigate the preventive effects of curcumin against acute pancreatitis (AP) induced by caerulein in mouse and to elucidate possible mechanism of curcumin action.
Methods:
Curcumin (50 mg/kg/day) was intraperitoneally injected to Kun Ming male mice for 6 days, followed by injection of caerulein to induce AP. GW9662 (0.3 mg/kg), a specific peroxisome proliferator-activated receptor gamma (PPARγ) antagonist, was intravenously injected along with curcumin. Murine macrophage RAW264.7 cells were treated with 100 μmol/l curcumin for 2 h, and then stimulated with 0.1 μ g/ml lipopolysaccharide (LPS). Serum amylase and transaminase levels were measured at 10 h after AP. TNF-α level in mouse serum and cell culture medium were detected by ELISA. Expression of PPARγ and NF-κB were analyzed by RT-PCR and Western blot.
Results:
Curcumin significantly decreased the pancreas injury and reversed the elevation of serum amylase, ALT and AST activities and TNF-α level in mice with AP. Curcumin treatment inhibited the elevation of NF-κB-p65 in the nucleus of mouse pancreas AP group and RAW264.7 cells, but significantly increased the expression of PPARγ. GW9662 could abolish the effects of curcumin on serum levels of amylase, ALT, AST, TNF-α, and NF-κB level.
Interpretation & conclusions:
Our results suggest that curcumin could attenuate pancreas tissue and other organ injury by inhibiting the release of inflammatory cytokine TNF-α. These effects may involve upregulation of PPARγ and subsequent downregulation of NF-κB.
Keywords
Acute pancreatitis (AP)
curcumin
NF-κB
PPARγ
TNF-α

Acute pancreatitis (AP) is a common cause of emergency hospital admission, with an increase in the incidence rate during the past 30 years1–4. Studies have demonstrated that inflammatory cytokines play an important role in the induction and severity of AP5. Clinical and experimental studies have shown that serum tumour necrosis factor-α (TNF-α) and some interleukins (IL) are increased in patients with AP6–8. These inflammatory cytokines expressed by activated macrophages, locally increase the severity of AP9. A key regulator of cytokine induction is nuclear factor-κB (NF-κB), which is usually elevated in the pancreas during AP. Inhibition of NF-κB activation results in a decrease in the release of inflammatory cytokines7.
Curcumin, a yellow coloured phenolic pigment extracted from the rhizome of herb Curcuma longa, is widely reported to have potent anti-oxidative, anti-inflammatory and anti-carcinogenic effects. The anti-inflammatory action of curcumin seems to be closely related to inhibition of TNF-α and other inflammatory cytokines production and suppression of NF-κB activation by blocking phosphorylation of inhibitory factor I-kappa B kinase (IκB)10–12.
Peroxisome proliferator-activated receptor gamma (PPARγ), a ligand-activated transcriptional factor, plays an important role in inflammation. Studies have demonstrated that ligand activation of PPARγ inhibits the activity of NF-κB, and negatively regulates inflammatory responses in vivo1314. A study demonstrated that curcumin can activate PPARγ and thus may be used as an agonist of PPARγ15. Therefore, the present study was focused on the preventive effect of curcumin in AP in mouse model and to investigate the possible mechanism of action.
Material & Methods
Reagents: Curcumin, caerulein, PPARγ antagonist GW9662 and LPS were purchased from Sigma (USA). TRIzol reagent was purchased from Invitrogen (USA). The reverse transcription kit, Taq DNA polymerase, and PCR kit were purchased from Fermentas (USA). Rat anti-PPARγ, anti-NF-κB-p65, anti-β-actin and anti-lamin B antibodies were purchased from Santa Cruz Biotechnology (USA), and Goat anti-mouse IgG from GenScript Company (Nanjing). The EasyECL assay kit and nuclear protein extraction kit were purchased from Biomed Company, China. The BCA kit was purchased from Galen Company, Hongkong. The ELISA kit specific for rat TNF-α was from Wuhan Boster Biological Technology, Ltd., China. DMEM medum was purchased from Gibco, USA and foetal bovine serum was purchased from Biochrom (Germany).
Animals: Four-week old Kun Ming male mice, weighing 30±2 g, were purchased from the Experimental Animal Center of Shandong University School of Medicine, Shandong, PR China. The animals were housed 6 per cage under standardized conditions (25±3°C, 12 h light/dark cycle, humidity 50±10%) with free access to pelleted food and tap water. All experiments were conducted in the Department of Biochemistry and Molecular Biology, Shandong University School of Medicine with the approval of the Institutional Experimental Animal Care and Use Committee of Shandong University.
Animal treatment procedures: A total of 40 healthy Kun Ming male mice were randomly divided into four groups and were fasted for 12 h with free access to water. AP was induced by intraperitoneal injection of 50 μg/kg bodoy wt. (bw) caerulein (dissolved in normal saline) once per hour, repeated 7 times16. In the negative control group, caerulein was replaced by normal saline. In curcumin group, prior to caerulein injection, mice were intraperitoneally injected with 50 mg/kg bw curcumin (dissolved in 10% DMSO)11 once a day for 6 consecutive days. To block PPARγ activity, a separate group of animals was treated with PPARγ antagonist, GW9662 (0.3 mg/kg bw) along with curcumin through intravenous injection prior to AP induction. Ten hours after AP induction17, the animals were killed and blood and tissue samples were collected.
Detection of serum amylase and transaminase activities: Blood was drawn from ophthalmic arteries, allowed to clot and centrifuged at 12000 g/min for 10 min. The serum was preserved at -80°C prior to analysis. The serum amylase, ALT and AST activities were analyzed by an automatic biochemical analyzer.
Pathological examination of mice pancreas: The mice were euthanized and pancreases were removed. The general morphologies of the pancreases were observed and recorded. Each pancreas was divided into two sections. One half was used to make paraffin-embedded sections, which were stained by hematoxylin and eosin (H & E) and pathologically examined. Schmidt criterion was employed for histopathological scoring in pathological evaluation18. The other half was subjected to RNA and protein extraction.
Curcumin treatment of murine macrophage RAW264.7 cells: Murine macrophage RAW264.7 cells (kindly provided by Prof. Chengjiang Gao from the Immunology Institute of Medical School of Shandong University) were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10 per cent foetal bovine serum, 50 units/ml penicillin and 50 μg/ml streptomycin at 37°C in a humidified CO2 incubator (5% CO2, 95% air). RAW 264.7 cells were plated at a density of 6×105/well in a 6-well plate. For all experiments, cells were grown to 80-90 per cent confluence. Cells were incubated in serum-free DMEM for 2 h and incubated with 100 μmol/l curcumin (dissolved in DMSO) at 37°C for another 2 h. After that, lipopolysaccharide (LPS) was added to achieve a final concentration of 0.1 μg/ml. Stimulation continued for 4 h, after which the supernatant was collected for TNF-α determination and cells were lysed for subsequent RNA or protein isolation. Triplicate samples were used for each treatment.
ELISA detection of TNF-α level: Enzyme-linked immunosorbent assays (ELISAs) were used to detect TNF-α level in mouse serum and the culture medium of RAW264.7 cells according to the manufacturer's instructions. All TNF-α determinations were performed in duplicate serial dilutions.
Reverse transcription-polymerase chain reaction (RT-PCR) for PPARγ mRNA expression in pancreases and RAW264.7 cells: Total RNAs in pancreases and RAW 264.7 cells were extracted with Trizol reagent according to the manufacturer's protocol and quantified by UV spectrophotometry. A total of 3 μg RNA was reverse transcribed into cDNA, and the resulting product was used as a template for PCR amplification of PPARγ genes. The PCR reaction system was set as 50 μl. β-actin was used as an internal reference under the same experimental conditions. Primers for PPARγ and β-actin were synthesized by Shanghai Sangon Biological Engineering Technology & Service Co., Ltd. Primers for PPARγ (477 bp) were: sense, 5’-CGTGATGGAAGACCACTCGC-3’, antisense, 5’- AACCTGATGGCATTGTGAGA-3’; for β-actin (218 bp) were: sense, 5′-TGGTGGGAATGGGTCAGA-3’, antisense, 5′-ACGGTTGGCCTTAGGGTT-3’. The cDNA was subjected to denaturation at 94°C for 2 min, followed by 32 cycles of 94°C for 30 sec, 56°C for 30 sec, and 72°C for 1 min, followed by a final extension step at 72°C for 10 min. PCR products were separated on 1 per cent agarose gel by electrophoresis, and the images were captured with the Gel Imaging System. Quantity One Software (? Source) was used to analyze the relative quantity of each band. The relative amount of PPARγ mRNA was normalized to β-actin.
Western blot analysis of NF-κB and PPARγ protein expression in pancreases and RAW264.7 cells: Total cellular and nuclear proteins in pancreas and RAW264.7cells were extracted and quantified with the BCA protein assay kit. Equal amounts of protein (50 μg) were separated by sodium dodecylsulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to a polyvinylidine difluoride (PVDF) membrane. The membrane was blocked for 1 h in TBS buffer (20mM Tris-HCl, 150mM NaCl, pH7.5) containing 5 per cent defatted milk at room temperature. The membrane was then incubated with primary antibody (mouse antihuman NF-κB-p65, PPARγ, β-actin or lamin B antibody; diluted at 1:1000) at room temperature for 2 h and at 4°C overnight. After washing in TBS buffer, the membrane was incubated with a HRP-coupled secondary antibody diluted 1:2000 for 1 h. Membrane-bound horse radish peroxidase-labelled protein bands were detected with enhanced chemiluminescent reagents and chemiluminescent signals were detected using X-ray film. The relative amount of each band was analyzed with Quantity One software. The ratios of PPARγ:β-actin and nuclear NF-κB:lamin B were employed to express the relative amounts of both proteins.
Statistical analysis: The data were analyzed with SPSS 10.0 software (SPSS Co., II, USA). Inter-group comparisons were performed with one-way ANOVAs. P<0.05 was considered as statistically significant.
Results
Pathological examination of mouse pancreas: Compared with the control group (histopathological scoring: 1.5±0.2, Fig. 1A), the mouse pancreas in the AP group (histopathological scoring: 13.2±1.7, Fig. 1B) showed haemorrhage and exudation. The pancreatic acinuses were diffusely expanded, the pancreatic cells underwent necrosis, and the inter-lobular and perivascular regions were infiltrated with multiple leukocytes. The necrosis rate and inflammatory cell infiltration were significantly (P<0.05) decreased in the pancreas in the curcumin group (histopathological scoring: 6.9±0.8, Fig. 1C) as compared to the AP group.
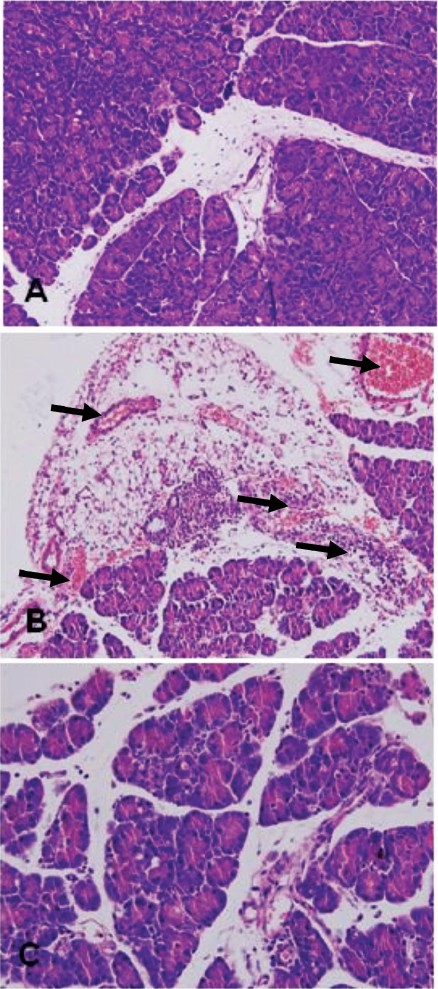
- Pathological changes in the mouse pancreas (HE staining, 200X). A: Pancreatic histopathology in control group. B: Pancreatic histopathology in AP group. C: Pancreatic histopathology in curcumin group.
Effect of curcumin on serum amylase and transaminase activities: Five hours after induction of AP, mice in the control group were still active, whereas mice in AP group were sick and reduced their physical activities. Ten hours after induction of AP, the serum amylase, ALT and AST activities of mice in the AP group were increased in comparison with those in the control group (P<0.01, Table). However, curcumin treatment significantly reversed the elevation of serum amylase, ALT and AST activities in AP mice (P<0.05, Table).

Effect of curcumin on the release of TNF-α: In the in vivo experiment, mouse serum TNF-α level increased in the AP group at 10 h after induction of AP (P<0.05) compared to control, whereas curcumin treatment significantly reduced the serum TNF-α level in AP mice (P<0.05) (Table). TNF-α levels in the culture media of RAW264.7 cells were significantly elevated after LPS stimulation (P<0.05) compared to control which significantly reduced in the presence of curcumin (P<0.05).
Effect of curcumin on nuclear NF-κB expression: Western blotting revealed that expression of NF-κB-p65 was significantly (P<0.05) increased in the nucleus of pancreas in AP group compared to the control group. Curcumin treatment inhibited the elevation of NF-κB-p65 in AP mice (Fig. 2). After LPS stimulation, NF-κB expression was significantly increased (P <0.05) in RAW264.7 cells and considerably decreased in the presence of 100 μmol/l curcumin (Fig. 3).
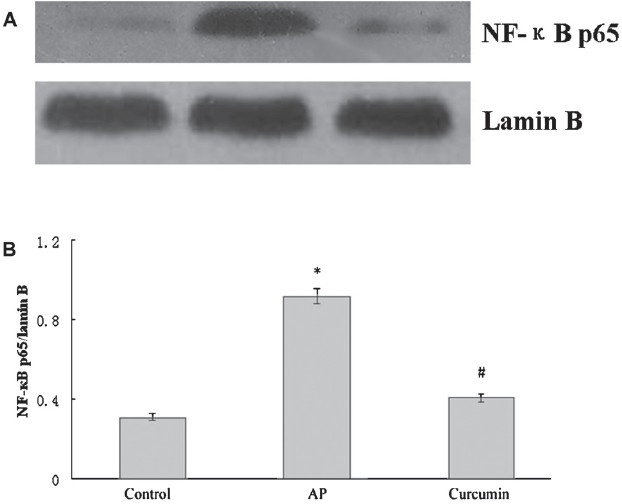
- Effect of curcumin on nuclear NF-κB expression in the pancreas of AP mice. (A) Western blot analysis of nuclear NF-κB expressionin in pancreas. (B) The graph shows densitometric analysis of the NF-κB relative to lamin B. *P<0.05 vs control group; #P<0.05 vs AP group. Values are mean±SD of 10 mice/group.
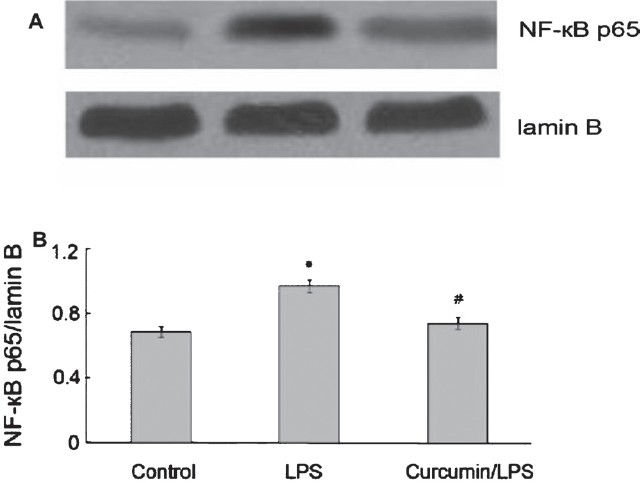
- Effect of curcumin on NF-κB expression in RAW264.7 cells. (A) Western blot analysis of NF-κB expressionin in RAW264.7 cells. (B) The graph shows densitometric analysis of the NF-κB relative to lamin B. *P <0.05 vs control group; #P<0.05 vs LPS group. Values are mean±SD (n=3).
Effect of curcumin on PPARγ expression: PPARγ expression was reduced at both the mRNA and protein levels in the pancreas of AP group, and this effect was significantly (P<0.05) reversed by curcumin treatment (Fig. 4 A and B). Similarly, after LPS (0.1 μg/ml) stimulation, PPARγ was decreased at both the mRNA and protein levels in RAW264.7 cells and was significantly (P<0.05) increased compared to control at both levels in the presence of 100 μmol/l curcumin (Fig. 5 A and B).
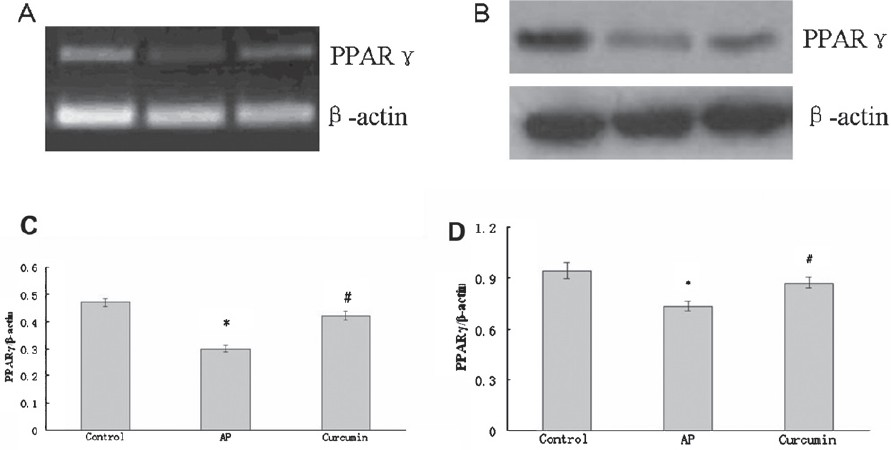
- Effect of curcumin on PPARγ expression in the pancreas of AP mice. (A) RT-PCR analysis of PPARγ mRNA expression in the pancreas. (C) The graph shows densitometric analysis of PPARγ relative to β-actin. *P<0.05 vs control group; #P<0.05 vs AP group. (B) Western blot analysis of PPARγ protein expression in the pancreas. (D) The graph shows densitometric analysis of PPARγ relative to β-actin. *P<0.05 vs control group; #P<0.05 vs AP group. Values are mean±SD of 10 mice / group.
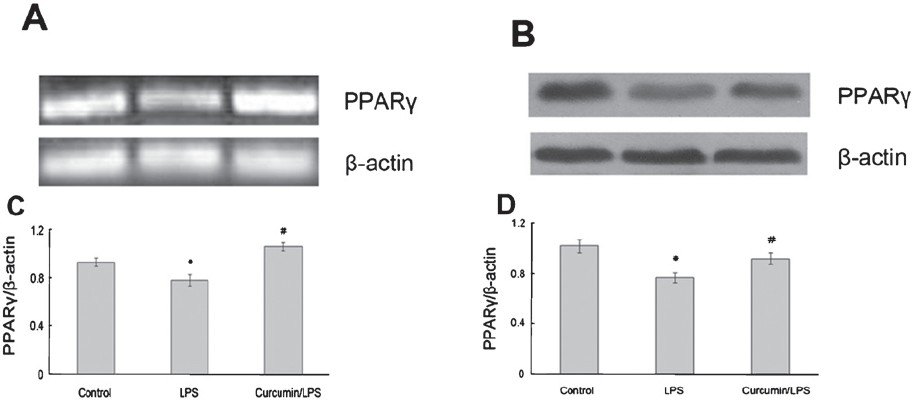
- Effect of curcumin on PPARγ expression in RAW264.7 cells. (A) RT-PCR analysis of PPARγ mRNA expression in RAW264.7 cells. (C) The graph shows densitometric analysis of PPARγ relative to β-actin. *P<0.05 vs control group; #P<0.05 vs LPS group. (B) Western blot analysis of PPARγ protein expression in RAW264.7 cells. (D) The graph shows densitometric analysis of PPARγ relative to β-actin. *P<0.05 vs control group; #P<0.05 vs LPS group. Values are mean±SD of (n=3).
Effect of the PPARγ antagonist GW9662: GW9662 partially counteracted the effects of curcumin on serum amylase, ALT, AST and TNF-α in AP mice (Table). A Western blot analysis of NF-κB expression showed that curcumin-mediated downregulation of nuclear NF-κB-p65 protein was significantly (P <0.05) suppressed by GW9662 (Fig. 6).
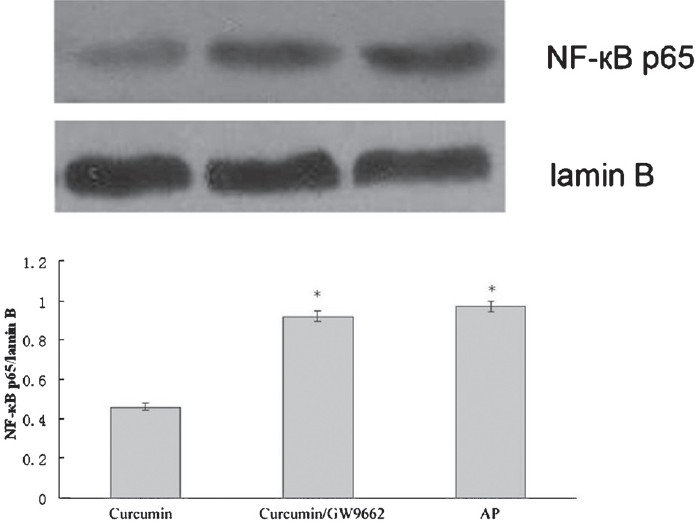
- Effect of curcumin plus GW9662 on nuclear NF-κB-p65 protein levels in the pancreas. *P<0.05 vs curcumin group.
Discussion
Curcumin has been shown to suppress multiple signaling pathways by interacting with a wide variety of proteins and modify their expression and activity, including inflammatory cytokines and enzymes, transcription factors, and gene products linked with cell survival, proliferation, invasion, and angiogenesis19. The aim of the present study was to investigate the potential preventive effect of curcumin to AP generation by using caerulein-induced pancreatitis in mouse model. Our results demonstrated that intraperitoneal injection of caerulein caused necrosis and haemorrhage and neutrophils infiltration in the mouse pancreas in AP group. Serum amylase and transaminase activities and TNF-α levels were all significantly elevated in the AP group. Curcumin treatment significantly decreased the histopathological score and reversed the elevation of serum amylase, ALT and AST activities and TNF-α levels. Pancreatic cell injury and enzyme release induce activation of pro-inflammatory cells, thereby resulting in the production of a large number of inflammatory mediators, which cause a systemic inflammatory response and multiple organ injury2021. Our results showed that curcumin not only attenuated pancreatic tissue injury, but also attenuated other organ injury by inhibiting the release of inflammatory cytokine TNF-α. Consistent with the in vivo experiment, TNF-α levels in the culture media of RAW264.7 cells were significantly elevated after LPS stimulation, while being significantly reduced in the presence of curcumin. Previous studies showed that curcumin has profound effects on modulation of TNF-induced signaling, as well as inhibition of expression of TNF. Curcumin treatment inhibited LPS or phorbol methyl acetate (PMA)-induced TNF-α levels in dendritic cells, macrophages, monocytes alveolar macrophages, and endothelial and bone marrow cells19. Curcumin inhibited the expression of TNF mRNA in the livers of copper uploaded rats and CCl4-induced hepatic fibrosis22. Our results were in agreement with Gukovsky et al11 who demonstrated that curcumin prominently reduced tissue injury as well as cytokine damage in both cerulein- and ethanol-induced pancreatitis, and curcumin's inhibitory effect on inflammatory mediators resulted in improvement in disease severity as measured by histology, serum amylase, pancreatic trypsin, and neutrophil infiltration. Gulcubuk et al23 demonstrated that curcumin failed in the reduction of tissue injury, and suggested that the failure might have been associated with the type of the experimental model used and the observation period and the level of cytokine.
The mechanism by which curcumin induces its anti-inflammatory effects is yet to be elucidated. Activation of transcription factor NF-κB, a key regulator of inflammatory molecule expression, is an early event in experimental pancreatitis and correlates with the inflammatory response. The present study revealed that expression of NF-κB-p65 was significantly increased in the nucleus of mouse pancreas in AP group and curcumin treatment inhibited the elevation of NF-κB-p65. After LPS stimulation, NF-κB expression was significantly increased in RAW264.7 cells and considerably decreased in the presence of curcumin. Samuhasaneeto et al24 suggested that curcumin treatment improved liver histopathology in early stage of ethanol-induced liver injury by reduction of oxidative stress and inhibition of NF-κB activation. Gukovsky et al11 reported that curcumin decreased inflammation by markedly decreasing activation of NF-κB and activator protein-1 (AP-1) as well as inhibiting mRNA induction of IL-6, TNF-α, and inducible nitric oxide synthetase (iNOS) in the pancreas in two rat models of experimentally-induced pancreatitis, assessed by DNA binding and degradation of inhibitory I-κB proteins. They indicated that blocking key signals of the inflammatory response ameliorates pancreatitis in both ethanol and nonethanol models8. Under normal physiological circumstances, inactivated NF-κB is sequestered by the inhibitor I-κB in the cytoplasm. When cells are stimulated by inflammatory factors, I-κB is phosphorylated and degradated through the ubiquitin-proteasome system. Then NF-κB is activated and translocates to the nucleus to initiate transcription of TNF-α and multiple pro-inflammatory factors25. Curcumin completely blocks TNF-induced NF-κB activation through the inhibition of phosphorylation of NF-κB, leading to suppression of cell survival, proliferative and inflammatory gene products26. Other studies showed that curcumin inhibited TNF-induced NF-κB activation by inhibiting proteasomal activities instead of IκBα kinase (IKK) activation in MCF-7 cells27. Our results indicated that elevation and activation of NF-κB might be a key process in the pathogenesis of AP, and that inhibition of NF-κB by curcumin should be explored as a preventive and therapeutic means of AP control and treatment.
Previous studies reported that curcumin significantly inhibited cell proliferation, induced apoptosis, inhibited of hepatic stellate cells (HSC) activation and suppressed hepatic fibrogenesis, attenuated STZ-induced memory deficits, eliminated ox-LDL stimulated HSC activation by stimulating PPAR-γ activity. Blocking the trans-activity by PPAR-γ antagonist significantly decreased these effects of curcumin2628–31. Studies performed by Jacob et al32 suggested that the anti-inflammatory effect of curcumin is mediated by the upregulation of PPAR-γ activation. Combined usage of GW9662 significantly impaired the function of curcumin in an experimental model of sepsis33. The present study showed that PPARγ expression was reduced at both the mRNA and protein levels in AP group mouse pancreas and in LPS-stimulated RAW264.7 cells, and curcumin treatment significantly increased the expression of PPARγ. Activation of PPAR-γ suppressed inflammatory responses by inhibiting the release of IL-1β, IL-6, TNF-α and expression of NF-κB and the adhesion molecule ICAM-13435. Concurrent administration of curcumin and GW9662 in the mouse with acute pancretitis abolished the effects of curcumin on serum levels of amylase, the liver enzymes ALT and AST, and TNF-α. A Western blot analysis of NF-κB expression showed that curcumin-mediated downregulation of nuclear NF-κB-p65 protein was significantly suppressed by GW9662, which suggests that downregulation of NF-κB by curcumin is mediated through PPAR-γ. Investigation conducted by Ghosh et al 36in Sprague-Dawley rats with chronic renal failure suggested that curcumin in a dose dependent manner antagonized the TNF-α-mediated decrease in PPAR-γ and blocked transactivation of NF-κB and repression of PPARγ. Collectively, these results suggest that the preventive effects of curcumin against AP were associated with PPAR-γ upregulation and subsequent NF-κB downregulation, indicating an important role of curcumin as a PPAR-γ agonist.
In summary, this study showed that curcumin induced anti-inflammatory effects caused by the upregulation of PPAR-γ were associated with the NF-κB pathway. Future studies are warranted to confirm this conclusion.
This work was supported by The Science Development Project of Shandong Province (2006GG2202021), China.
Competing interests: The authors declare that they have no competing interests.
References
- Time trends in incidence, etiology, and case fatality rate of the first attack of acute pancreatitis. Scand J Gastroenterol. 2011;46:1389-98.
- [Google Scholar]
- First attack of acute pancreatitis in Sweden 1988 - 2003: incidence, aetiological classification, procedures and mortality - a register study. BMC Gastroenterol. 2009;9:18.
- [Google Scholar]
- Trends in the epidemiology of the first attack of acute pancreatitis: a systematic review. Pancreas. 2006;33:323-30.
- [Google Scholar]
- Epidemiology, aetiology and outcome of acute and chronic pancreatitis: An update. Best Pract Res Clin Gastroenterol. 2008;22:45-63.
- [Google Scholar]
- The pathogenesis of L-arginine-induced acute necrotizing pancreatitis: inflammatory mediators and endogenous cholecystokinin. J Physiol Paris. 2000;94:43-50.
- [Google Scholar]
- Proinflammatory cytokines in early assessment of the prognosis of acute pancreatitis. Am J Gastroenterol. 1999;94:213-8.
- [Google Scholar]
- Suppression of NF-kappaB activation and cytokine production by N-acetylcysteine in pancreatic acinar cells. Free Radic Biol Med. 2000;29:674-83.
- [Google Scholar]
- Profiles of pro-inflammatory cytokines in the serum of rabbits after experimentally induced acute pancreatitis. Cytokine. 2002;17:53-9.
- [Google Scholar]
- Macrophage ablation attenuates adenoviral vector-induced pancreatitis. Surgery. 2005;137:545-51.
- [Google Scholar]
- Curcumin inhibition of inflammatory cytokine production by human peripheral blood monocytes and alveolar macrophages. Pharmacol Res. 1999;39:41-7.
- [Google Scholar]
- Curcumin ameliorates ethanol and nonethanol experimental pancreatitis. Am J Physiol Gastrointest Liver Physiol. 2003;284:G85-95.
- [Google Scholar]
- Curcumin suppresses Janus kinase-STAT inflammatory signaling through activation of Src homology 2 domain-containing tyrosine phosphatase 2 in brain microglia. J Immunol. 2003;171:6072-9.
- [Google Scholar]
- Phosphorylation of PPARgamma via active ERK1/2 leads to its physical association with p65 and inhibition of NF-kappabeta. J Cell Biochem. 2003;90:732-44.
- [Google Scholar]
- Curcumin potentiates antitumor activity of gemcitabine in an orthotopic model of pancreatic cancer through suppression of proliferation, angiogenesis, and inhibition of nuclear factor-kappaâ-regulated gene products. Cancer Res. 2007;67:3853-61.
- [Google Scholar]
- Activation of peroxisome proliferator-activated receptor-gamma contributes to the inhibitory effects of curcumin on rat hepatic stellate cell growth. Am J Physiol Gastrointest Liver Physiol. 2003;285:G20-30.
- [Google Scholar]
- Caerulein-induced acute necrotizing pancreatitis in mice: Protective effects of proglumide, benzotript, and secretin. Gastroenterology. 1985;88:1192-204.
- [Google Scholar]
- Recombinant human interleukin-11 decreases severity of acute necrotizing pancreatitis in mice. Pancreas. 2000;21:134-40.
- [Google Scholar]
- Morphometric characteristics and homogeneity of a new model of acute pancreatitis in the rat. Int J Pancreatol. 1992;12:41-51.
- [Google Scholar]
- Curcumin inhibits proliferation,invasion, angiogenesis and metastasis of different cancers through interaction with multiple cell signaling proteins. Cancer Lett. 2008;269:199-225.
- [Google Scholar]
- Acute pancreatitis and organ failure: pathophysiology, natural history, and management strategies. Curr Gastroenterol Rep. 2004;6:99-103.
- [Google Scholar]
- Role of cytokines and oxidative stress in the pathophysiology of acute pancreatitis: therapeutical implications. Curr Drug Targets Inflamm Allergy. 2002;1:393-403.
- [Google Scholar]
- Curcumin protects the rat liver from CCl4-caused injury and fibrogenesis by attenuating oxidative stress and suppressing inflammation. Mol Pharmacol. 2008;73:399-409.
- [Google Scholar]
- Effects of curcumin on tumour necrosis factor-alpha and interleukin-6 in the late phase of experimental acute pancreatitis. J Vet Med A Physiol Pathol Clin Med. 2006;53:49-54.
- [Google Scholar]
- Curcumin decreased oxidative stress, inhibited NF-kappaB activation, and improved liver pathology in ethanol-induced liver injury in rats. J Biomed Biotechnol. 2009;2009:981963.
- [Google Scholar]
- Curcumin: Biological and medicinal properties. In: Ravindran PN, Nirmal Babu K, Shivaraman, eds. Turmeric: the genus Curcuma. Boca Raton FL: The CRC Press; 2006. p. :297-368.
- [Google Scholar]
- Effect of selected phytochemicals and apple extracts on NF-kappaB activation in human breast cancer MCF-7 cells. J Agric Food Chem. 2007;55:3167-73.
- [Google Scholar]
- The molecular targets and therapeutic uses of curcumin in health and disease. Adv Exp Med Biol. 2007;595:1-512.
- [Google Scholar]
- Activation of peroxisome proliferator-activated receptor-gamma by curcumin blocks the signaling pathways for PDGF and EGF in hepatic stellate cells. Lab Invest. 2008;88:529-40.
- [Google Scholar]
- Involvement of PPAR-gamma in curcumin-mediated beneficial effects in experimental dementia. Naunyn Schmiedebergs Arch Pharmacol. 2010;381:529-39.
- [Google Scholar]
- Curcumin eliminates oxidized LDL roles in activating hepatic stellate cells by suppressing gene expression of lectin-like oxidized LDL receptor-1. Lab Invest. 2009;89:1275-90.
- [Google Scholar]
- Mechanism of the antiinflammatory effect of curcumin: PPAR-γ activation. PPAR Res 2007. 2007;2007:89369.
- [Google Scholar]
- The anti-inflammatory effect of curcumin in an experimental model of sepsis is mediated by up-regulation of peroxisome proliferator-activated receptor-gamma. Crit Care Med. 2006;34:1874-82.
- [Google Scholar]
- A novel therapy for colitis utilizing PPAR-gamma ligands to inhibit the epithelial inflammatory response. J Clin Invest. 1999;104:383-9.
- [Google Scholar]
- Peroxisome proliferator-activated receptors and inflammation: from basic science to clinical applications. Int J Obes Relat Metab Disord. 2003;27(Suppl 3):S41-5.
- [Google Scholar]
- Curcumin ameliorates renal failure in 5/6 nephrectomized rats: role of inflammation. Am J Physiol Renal Physiol. 2009;296:F1146-57.
- [Google Scholar]




