
Translate this page into:
Adhesion molecules facilitate host-pathogen interaction & mediate Mycobacterium tuberculosis pathogenesis
For correspondence: Dr Laxman S. Meena, CSIR-Institute of Genomics & Integrative Biology, Mall Road, Delhi 110 007, India e-mail: meena@igib.res.in
-
Received: ,
This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
This article was originally published by Wolters Kluwer - Medknow and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Most of the microorganisms display adhesion molecules on their surface which help them to bind and interact with the host cell during infection. Adhesion molecules help mycobacteria to colonize and invade immune system of the host, and also trigger immune response explicated by the host against the infection. Hence, understanding the signalling pathways illustrated by these molecules to enhance our knowledge on mycobacterial survival and persistence inside the host cell is required. Hence, this review was focussed on the role of adhesion molecules and their receptor molecules. The various mechanisms adopted by adhesion molecules to bind with the specific receptors on the host cell and their role in invasion and persistence of mycobacterium inside the host cell are explained.
Keywords
Adhesion
fibronectin
fibronectin-binding protein
lipoarabinomannan
macrophages
Mycobacterium tuberculosis
proline glutamic polymorphic CG repetitive sequence

Introduction
Tuberculosis (TB) remains a worldwide cause of increasing morbidity and mortality despite major advances in anti-TB drug administration and treatment12. Increasing cases of multidrug resistance and extensive drug resistance with co-infection of HIV has further added to the prevalence of the active disease3. According to the WHO Report 2017, 6.3 million new TB cases were reported in 20164. However, TB motility rate fell by 37 per cent in between 2000 to 2016. In spite of that, the Report has shown an increasing prevalence of the disease worldwide4.
An understanding of interaction between host cell surface receptors with pathogen's surface-associated adhesion molecules is required to gain access to the mechanism involved in host-pathogen interactions. In reference to this, the bacterial strategy to countervail the host defence and how it persists for a longer time within the host can be further analyzed56. Identification of functioning and unified system of molecules present in host pathogenesis is well identified through studying protein-protein interactions, which contributes to cellular mechanism. On these grounds, it would be interesting to elaborate functions of adhesion molecules which are majorly involved in mediating interaction between the pathogen and the host cell5678 (Fig. 1 and Table). Adhesion molecules are cell surface molecules, which bind to receptors or with soluble macromolecules present in extracellular membrane of host to promote cell adhesion and are involved in host pathogenesis78. Several adhesion molecules found in mycobacteria such as fibronectin-binding proteins (FnBPs) and heparin-binding haemagglutinin (HBHA), which are involved in adherence and promote internalization of the Mycobacterium tuberculosis into the host cells, facilitate bacterial colonization9. Interaction of these adhesions with host cell surface receptors not only helps in attachment and invasion but further exhibiting a cascade of signalling such as interferon (IFN)-γ response and activation of mitogen-activated protein kinases (MAPKs) pathway, which promotes pro- and/or anti-inflammatory events by stimulating an immune response8. Further, adhesion molecules not only trigger the immune response but also interfere with the host signalling and modulate its intracellular mechanism (Fig. 2). Hence, exploiting the role of adhesion molecules is important to understand host pathogenesis inducted by M. tuberculosis.
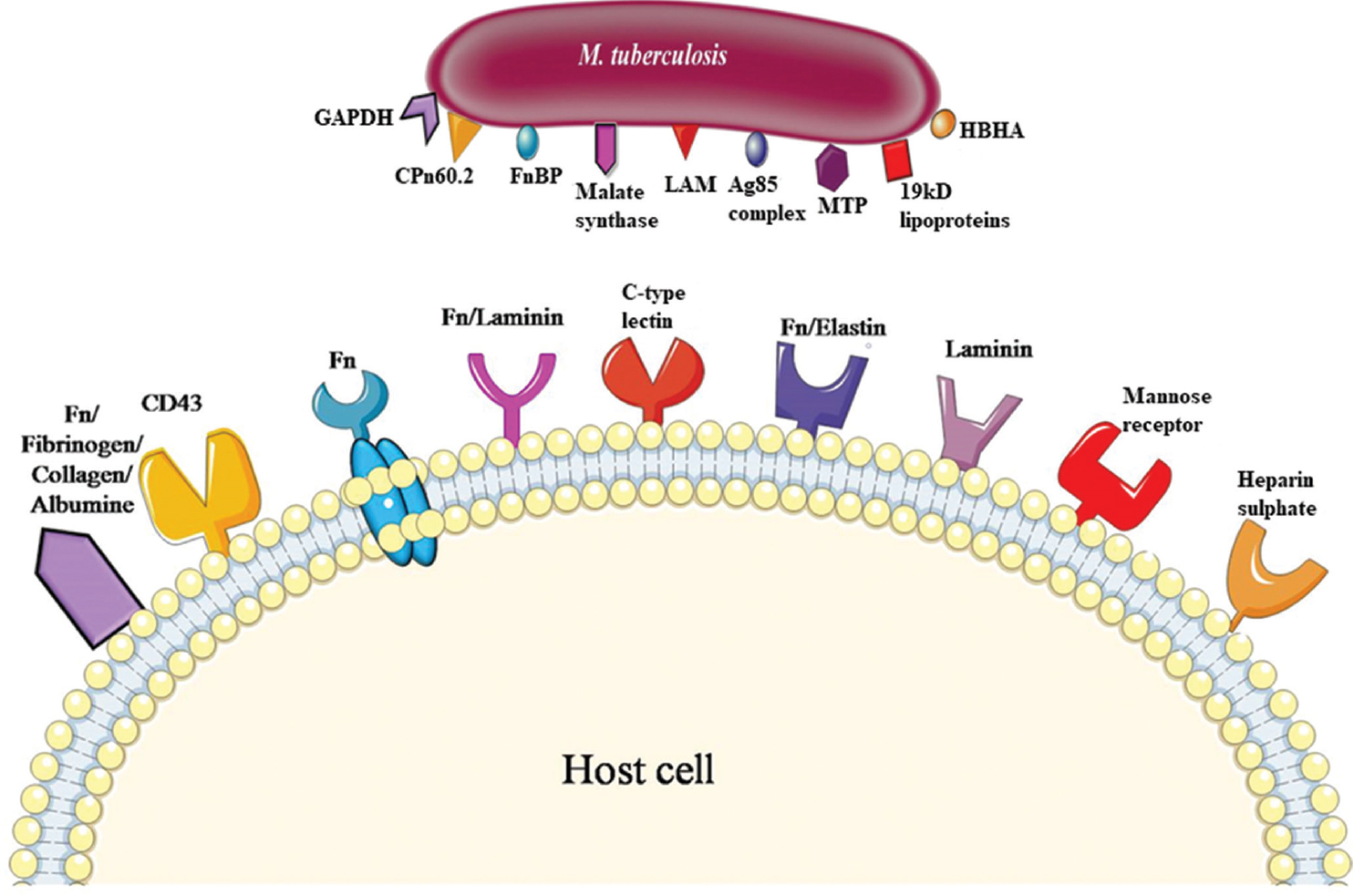
- Illustrative representation of mycobacterial adhesions and their respective receptors on host cell (Structures portrayed here do not necessarily imitate the actual receptor structure). GAPDH, glyceraldehyde 3-phosphate dehydrogenase; CPn60.2, chaperonin 60.2; FnBP, fibronectin binding protein; LAM, lipoarabinomannan; Ag 85, antigen 85 complex; MTP, Mycobacterium tuberculosis pili; HBHA, heparin-binding hemagglutinin A; Fn, fibronectin.
| Adhesins present in pathogen’s surface | Receptors present in host cell | Pathogenesis | References |
|---|---|---|---|
| Man-LAM | TLRs, C-type lectin | Interference with calcium signalling, blocks signalling of DC | 6151920 |
| FnBp | Fn | Reorganization of actin cytoskeleton, stimulates coronin/TACO which inhibits phagosome lysosome fusion | 2228 |
| Cpn60.2 | CD43 | Stimulates production of TNFα, interferes in antigen presentation | 2931 |
| HBHA | Sulphated surface receptors such as heparin sulphate | Mycobacterial dissemination | 333738394042 |
| Antigen 85 complex | Fn | Maintains cell wall integrity of mycobacteria | 454649 |
| 19 kDa protein | Mannose receptor | Decreases antigen presentation by inhibiting HLA-DR protein | 5357 |
| Malate synthase | Laminin, Fn, A549 lung epithelial cells | Helps mycobacteria to survive under hypoxic condition | 5960 |
| MTP | Laminin, epithelial cells | Mainly promotes adhesion | 6667 |
| GAPDH | Fn, fibrinogen, albumin, collagen | Helps to uptake iron from host cell | 6873 |
Man-LAM, mannose lipoarabinomannan; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; MTP, Mycobacterium tuberculosis pili; HLA-DR, human leukocyte antigen-antigen D-related; HBHA, heparin binding haemagglutinin; TACO, tryptophan aspartate containing coat protein; TLRs, Toll-like receptors; Fn, fibronectin; FnBp, fibronectin binding protein; TNFα, tumour necrosis factor alpha; DC, dendritic cells
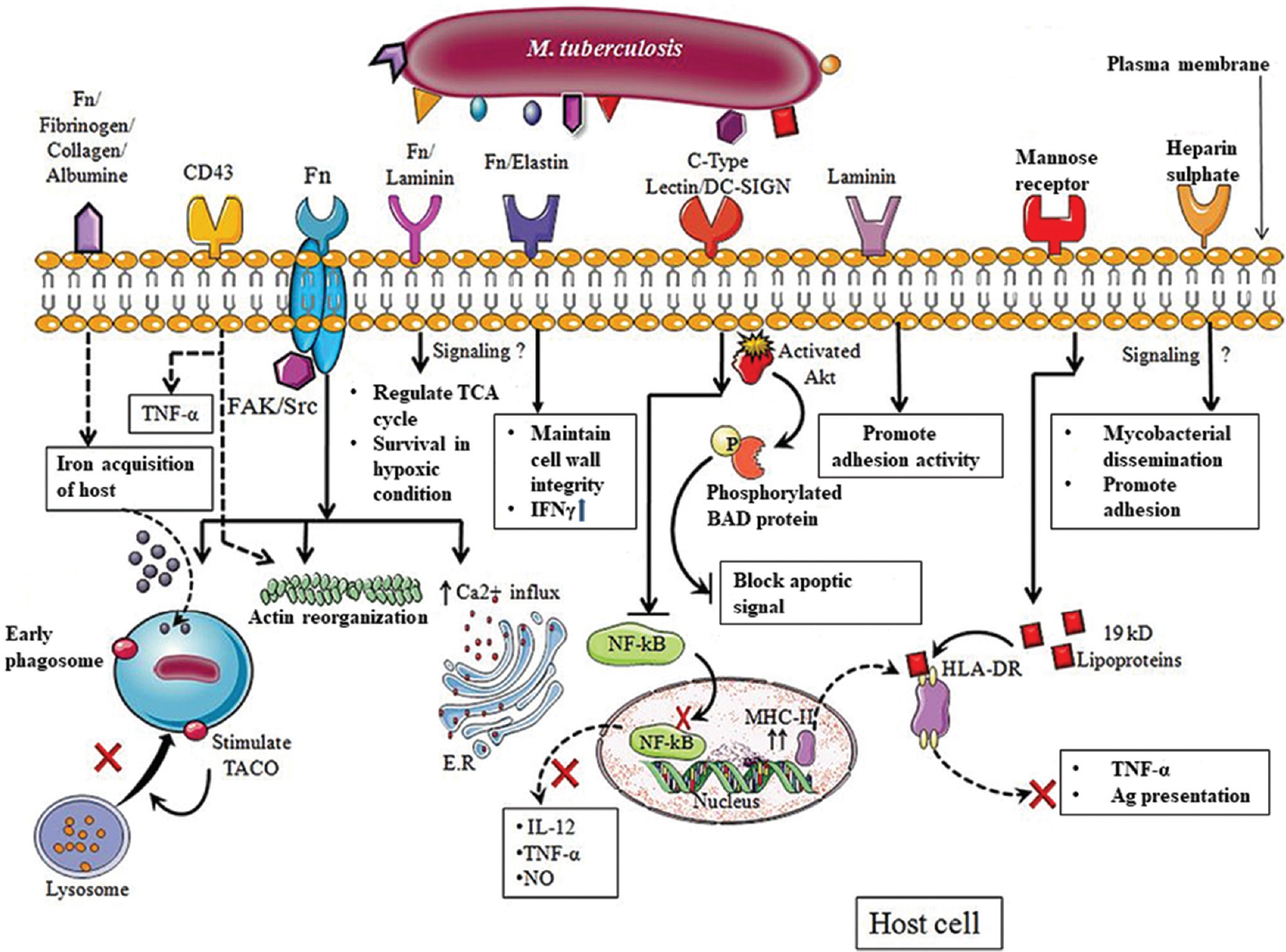
- Schematic representation of signalling induced by adhesion molecules inside the host cell. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) binding to fibrinogen (Fn) on the host cell induces the uptake of iron which is to be acquired by the host cell for its own use. Cpn60.2 binding to CD43 induces interferon-γ (IFN-γ) production and also leads to actin reorganization. Adhesion of fibronectin-binding protein (FnBP) to fibronectin stimulates FAK/Src kinase which also leads to actin reorganization and also recruits tryptophan aspartate containing coat protein to early phagosome and prevents its fusion with lysosome. It also triggers calcium upregulation. Malate synthase binding with fibronectin or laminin has been shown. It mainly helps bacteria to survive under hypoxic conditions inside the host cell. Antigen-85 complex binding with fibronectin/elastin induces interferon-γ (IFN-γ) which indicates that it also participates in host cell signalling mechanism. Binding of lipoarabinomannan to C-type lectin/DC-SIGN stimulates Akt protein which phosphorylates Bad protein, and hence, the intrinsic apoptotic pathway is blocked. Mycobacterium tuberculosis pili binding to laminin promotes strong adhesion activity; 19 kDa protein binds with mannose receptor and blocks human leukocyte antigen-antigen D-related (HLA-DR) protein present on major histocompatibility complex II (MHC-II) and leads to delayed antigen presentation. Heparin-binding haemagglutinin binds to heparin present on host cell and triggers mycobacterial dissemination which is important for its survival inside the host cell.
Synergetic effect of adhesion and host cell signalling in TB infection
Lipoarabinomannan (LAM): Lipoarabinomannan (LAM) is a cell wall active polysaccharide of M. tuberculosis, which constitutes arabinose and mannose moieties. Multiple branched arabinofuranosyl (Ara) side chain modified with mannose residue encompasses LAM1011121314. Mannose-lipoarabinomannan (Man-LAM) is abundant in pathogenic mycobacteria such as M. tuberculosis and M. ulcerans while Ara-LAM is more excessive in non-pathogenic bacteria such as M. fortiutum and M. smegmatis1112. The phosphatidylinositol moiety in LAM helps the pathogen to anchor to the cell wall of the host cell14. It has been proposed that LAM, along with 13 lipomannans, is an immunomodulatory glycoconjugate molecule, which constitutes several chemical structures involved in M. tuberculosis disease progression151617. These structures have an ability to interact with their respective receptors on the host cell surface.
LAM interferes with calcium signalling: In the host cell an increase in the amount of calcium modifies calmodulin, which in turn activates Ca2+/Calmodulin-dependent protein kinase II (CaMKII)18. CaMKII is required to recruit early endosome antigen I (EEA1) to the phagosomal membrane which helps in vesicle fusion. LAM decreases Ca2+ concentration by either chelating Ca2+ or inhibiting CaMKII and phagosome maturation1519. Furthermore, the intracellular survival of mycobacteria has also been maintained by Man-LAM which takes part in blocking apoptotic signals. Man-LAM stimulates phosphorylation of apoptic protein Bad (pro-apoptotic protein), which in turn stimulates Akt protein kinase activity and interferes with apoptotic signals20. These illustrations clearly establish that Man-LAM helps mycobacteria to invade and persist for longer time inside the host cell and hence is involved in pathogenesis.
LAM blocks signalling of dendritic cells: In addition to macrophages, other cells such as dendritic cells (DCs) also display Toll-like receptors (TLRs) and C-type lectins on their surface21. TLRs are conserved receptors that can perceive pathogen-associated molecular patterns (PAMPs) and actuate innate as well as adaptive immunity against these pathogens. Man-LAM binds to C-type lectins which are involved in recognizing a wide variety of pathogens through their diverse carbohydrate structures that induce host-pathogen interaction. This also suggests that LAM interferes in lipopolysaccharide signalling exhibited by TLRs and inhibits production of interleukin (IL)-12 by DCs and suppresses the immune response620.
Fibronectin-binding protein (FnBP): It is a secretary adhesion protein on the mycobacterial surface, which proves to be a potent immunomodulator2223. FnBP complex constitutes antigen-85A, -85B, -85C (Ag85a, Ag85B, Ag85C) and FnBP A, B, C which play a crucial role in M. tuberculosis pathogenesis and also illustrate cell wall mycolyl transferase activity23. FnBPs mainly belong to proline glutamic polymorphic CG repetitive sequence family of genes present in mycobacteria242526. Previous studies have validated its binding to fibronectin (Fn), a multidomain glycoprotein on host surface22. Fn can act as a ligand to a number of integrin receptors. Hence, one can presume that the connection of FnBPs with integrin present on host surface is mediated by Fn receptor molecule. Moreover, in other bacterial species, such as Staphylococcus aureus, it has been established that Fn executes adhesion to live endothelial cells27 which leads to phagocytosis and activation of cytokines, i.e. a suitable immune response, is generated28 (Table). This supports the idea of Fn being an adhesion molecule involved in pathogenesis. FnBPs have been identified as members of microbial surface components recognizing adhesive matrix molecules in M. tuberculosis infection22. These are considered to constitute LPXTG motif in their C-terminal region which covalently attaches FnBp to the extracellular matrix (ECM)29. In other studies, it has also been demonstrated that Fn plays crucial role in matrix assembly. Fn binding leads to interaction of the virulent factor with integrin on the host cell, thus resulting in reorganization of actin cytoskeleton associated with tyrosine kinase activation. For example, stimulation of focal adhesion kinase (FAK) and Src kinases signalling has been shown to promote phoshpholipase activity which helps stimulate coronin/tryptophan aspartate-containing coat protein (TACO). TACO recruited to phagosomal surface will inhibit phagosomal maturation28. This exemplifies as to how Fn, an adhesion molecule, modulates host cell signalling and helps in survival of mycobacteria inside the granuloma for a longer period of time.
Cpn60.2: Molecular chaperones are proteins that play a vital role in maintaining cellular functions by facilitating protein folding. These are potentially involved in entry of mycobacteria inside the host cell and facilitate pathogenesis. There are two types of chaperone molecules found on M. tuberculosis cell surface, i.e. Cpn60.2 (also known as Hsp65) and Hsp70, which belongs to the family of heat shock proteins. Molecular chaperones can act like 'moonlighting proteins', suggesting that these can serve for varied functions30. Experiments have proved that almost 57 per cent of the bacterial association with macrophages is hindered by recombinant Cpn60.231. Hence, one can say that Cpn60.2 is a major adhesion molecule found on the surface of M. tuberculosis. Previous studies have proved that it helps in binding to the host cell through its attachment to a large sialylated glycoprotein present on the surface of the macrophage known as CD4331. Cpn60.2 plays a major role in cell survival mechanism inside the host and can act as a potent immunomodulator32. It has also been found that Cpn60.2 can activate human peripheral blood macrophages followed by stimulation and secretion of cytokines that brings B- and T-cell activation30. Furthermore, CD43 interaction with the pathogen regulates tumour necrosis factor (TNF)-α production by M. tuberculosis-infected macrophages cell33. Hence, CD43 is shown to be related with cell signalling events along with cytoskeleton rearrangement and other intracellular as well as extracellular functions30. According to evidences, Cpn60.2 has been unable to show anti-inflammatory activity34. Binding of Cpn60.2 with macrophages does not reveal enhanced expression of major histocompatibility complex II (MHC-II), Fc-γ receptors or any other free radicals. This suggests that these will interfere in antigen presentation and stimulation of T-lymphocytes, thus ensuring the survival of M. tuberculosis inside the host cell35. As a heat shock protein, Cpn60.2 is also involved in the survival of mycobacteria inside the host cell during stress conditions36. In M. leprae infection also, it plays a vital role in coordination with T-cells that help establish its role in M. tuberculosis. It stimulates monocyte cytokines which guides the formulation of TNF, IL-6 and IL-2, thus effectively mediating host immunity. It also constitutes highly conserved sequence due to which it is also involved in progression of autoimmune response.
Heparin-binding haemagglutinin (HBHA): This is a surface exposed 28 kDa molecule with 199 amino acid residues which can act as an adhesion molecule. Adherence property of mycobacteria to epithelial cells is specifically inhibited by heparin and other sulphated carbohydrates. This proves that HBHA is involved in adherence by binding to sulphated surface receptors37383940. It also acts as a methylated antigen that helps in HBHA-specific T-cell response41. According to Pethe et al42, HBHA disrupted mutant strain of BCG shows no difference in invading phagocytic cells in comparison to wild type, while these show 60 per cent reduction in adherence to pneumocytes. Hence, it is concluded that HBHA-mediated adherence is specific to non-phagocytic cells and that it is involved in mycobacterial dissemination which is essential for the pathogen to spread its infection3339. HBHA acts as a multifunctional adhesion molecule which is involved in auto-aggregation as well as bacterial-eukaryotic cell interaction39. Potential of HBHA in the detection of latent infection of TB has also been evaluated.
Antigen-85 (Ag85) complex: Ag85 complex serves as a major secretory product released by M. tuberculosis, which possesses Fn-binding property43. It constitutes three abundantly secreted proteins 85A (fbpA), 85B (fbpB) and 85C (fbC) encoded by Rv3804c, Rv1886c and Rv0129c, respectively, in M. tuberculosis44. These have been shown to be involved in disease pathogenesis as these stimulate uptake of M. tuberculosis via their interaction with Fn receptor molecules present on host cell4546. Armitige et al47 have demonstrated that disrupting genes of Ag85 complex from M. tuberculosis strains affects cell wall synthesis. Besides having role in cell wall biosynthesis, the surface Ag85 complex has also been found to exhibit cell wall mycolyl transferase activity during infection45.
Studies have shown that transfer of mycolylates is catalyzed by the mycolyl transferase48. It directs the formation of two products: α,α-trehalose monomycolate and α, α-trehalose dimycolate (a cord factor), which are useful in maintaining cell wall integrity in M. tuberculosis infection49. A significant proliferation and IFN-γ secretion in the peripheral blood leukocytes have been found to be induced by 32 kDa protein (Ag85) isolated from M. bovis50. Hence, it is suggested that Ag85 complex acts as a prominent marker in diagnosis of TB as it can be detected in blood and sputum of pulmonary TB patients51.
19 kDa protein: The 19 kDa lipoprotein is expressed by M. tuberculosis and other slow-growing pathogenic bacteria such as M. vaccae and not by fast-growing bacteria such as M. smegmatis52. It is a surface exposed glycolipoprotein, which acts as pathogen associated molecular patterns (PAMPs) that can be recognized by pattern recognition receptors such as TLRs. Hence, it is involved in the production of pro-inflammatory cytokines such as ILs and TNF-α, which further generate a significant immune response. It has been found to bind with mannose receptor present on host cell surface and promote adhesion of mycobacteria to host cell53. A structural motif of triacyl head group is attached to amino terminal cysteine which is expressed by these lipoproteins. Its interaction to host cell via TLR2 has been shown in previous studies54, indicating its participation in host cell signalling. In addition, Neufert et al55 have demonstrated that 19 kDa protein also acts as a priming agent in neutrophil activation. Further, these lipoproteins have been found to induce humoral as well as cellular immune responses by host cell towards M. tuberculosis which demonstrates its role in pathogenesis. It also confers TLR2-dependent inhibition of MHC-II expression and antigen processing56. Gehring et al57 have demonstrated that 19 kDa lipoprotein interferes in IFN-γ signalling by inhibiting IFN-γ-regulated human leukocyte antigen-antigen D-related (HLA-DR)57 protein which in turn inhibits IFN-γ induced expression of fcγ-R1 and decreases bacilli antigen processing and presentation on MHC-II to CD4+ T-cells. This evolves a mechanism for M. tuberculosis to survive and persist for a long time inside the cell. It is also described that this inhibitory effect shown by 19 kDa protein is restricted to MHC-II expression instead of MHC-I. In addition, 19 kDa protein also contributes in transport of nutrients through mycobacterial cell wall which helps in its survival. Ciaramella et al58 have reported that 19 kDa proteins majorly induce apoptosis in monocytes/macrophages by LpqH activation which in turn triggers TLR2 activation which upregulates the death receptor and signalling molecule, thus beginning death receptor signalling cascade. They demonstrated this using anti-19 kDa monoclonal antibodies and high amounts of M. tuberculosis H37Rv. This protein needs to be studied further to track mycobacterial strategy to invade and persist inside the host cell.
Malate synthase: Malate synthase is a multifunctional protein found in mycobacteria encoded by the gene glcB59. It is an extracellular protein which serves as an adhesin and has a significant role in host pathogenesis. This enzyme has been shown to bind with laminin, Fn and A549 lung epithelial cells, which is acquired by a unique C-terminal region60. As bacterial persistence inside the host undergoes low oxygen tension, the changes in malate synthase help bacteria to survive. The involvement of malate synthase in glyoxylate pathway has been also suggested. Assimilation of the carbon compound into tri carboxylic acid (TCA)61 cycle is accompanied by malate synthase, which can be utilized for the bacterial replication in macrophages62. Malate synthase serves to play vital role for growth of oxidizing co-factors involved in energy production and perseverance of carbon moieties during M. tuberculosis survival inside the host cells under hypoxic conditions59. It has also been demonstrated that malate synthase is recognized by serum antibodies easily; hence, it can be used in serodiagnostic assay63. Thus, malate synthase not only serves as an adhesion molecule but also helps bacteria to survive inside the host cell under stress conditions.
Mycobacterium tuberculosis pili (MTP): M. tuberculosis pili (MTP) are fine adhesion molecules, which belong to bacterial amyloid family called pili or curli. Using electron microscopy, Alteri et al64 have demonstrated that mycobacterium produces two distinct types of pili morphotypes, type IV and curli pili. In terms of adhesion property, MTPs have shown adherence properties that help them to bind with laminin present in the ECM of the host cell. Previously, it was also established that pili were involved in mediating specific recognition of receptors present in host cells64. Several studies have demonstrated that it is encoded by the gene, Rv3312A, which is further involved in tissue colonization and reported as potential virulence factor associated to M. tuberculosis pathogenesis64. Pili are also involved in bacterial aggregation, host cell interaction and biofilm formation which lead to colonization of mucosal surface. Ramsugit and Pillay65 demonstrated that MTP mutant-type mycobacterial strain was not able to adhere and invade THP-1 macrophages to that extent as compared to wild-type strain which further confirmed their adherence property65. They also found that MTP did not extensively influence the whole cytokine response of M. tuberculosis infected epithelial cells66. Alteri et al64, using immunoflourescence-based assay, analyzed the presence of interaction of MTP with A549 epithelial cells. Further, it was reported that adhesion between bacteria and epithelial cells was mediated by the hydrophobicity of pilian adhesion67. Earlier studies also show the role of MTP in biofilm formation. Hence, MTP is a major adhesion molecule which helps to invade the host cell and is involved in pathogenesis.
Glyceraldehyde 3-phosphate dehydrogenase (GAPDH): Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) is known to be a surface associated multifunctional protein6869. The adhesion property of the enzyme is exhibited towards Fn, fibrinogen, albumin and collagen68. It is also a vital moonlighting protein, i.e. it exhibits multiple functions69. It is demonstrated that the highly conserved enzyme converts glyceraldehyde-3-phosphate in the presence of inorganic phosphate and nicotinamide adenine dinucleotide (NAD+) to 1,3-bisphosphoglycerate. Meanwhile, NAD+ is reduced to NAD hydrogenase7071. Studies have shown that GAPDH is also involved in cellular signalling network; for example, phosphorylation of Siah-I by apoptosis signal regulating kinase I (ASKI). It is a stress kinase which triggers GAPDH-Siah-I stress signalling, thus suggesting that it is involved in stress signalling networks7072.
Uptake of iron from the host cell
Overexpression of GAPDH helps M. tuberculosis to uptake iron from host cells via its increased binding to transferrin, which is a host iron transport protein evolved during infection73. Past demonstration suggests that iron is important for persistence of the pathogen inside the host. Transferrin bound iron is utilized by transferrin receptor-1 (TFR-1) expressed in macrophages. Mycobacterium avium has been studied to interfere with TFR-1 and utilize transferrin bound protein for its own use which helps us to hypothesize similar roles in M. tuberculosis pathogenesis. GAPDH has also been reported to be involved in apoptosis induction and transcription regulation74. Hence, by targeting GAPDH and signalling cascade, we could probably open up distinct channels for the evaluation of novel TB drugs to eradicate tuberculosis.
Conclusion
The pathogenic effects of adhesion molecules in TB pathogenesis and its intimate relation with host cellular signalling require more researches in this direction to be clearly interpreted. According to previous studies, the current status is that the factual role of these adhesion molecules remains elusive. Adhesion molecules are the initiation mediators of mycobacterial interaction with the host cell. Hence, gaining access to the overall structural and functional aspects of adhesion molecules involved in host-pathogen interaction is essential. Further elucidation of their importance in host signalling can be accessed via methods such as RNA interference and knock-out techniques which can find their actual role and mechanism of action inside the host cell. The involvement of a cooperative mechanism in host invasion can also be studied. This information is essential in targeting these adhesion molecules to develop new TB vaccines and treatment strategies, which can help to cope up with the epidemiology of the disease and thus fulfil our ultimate goal to control this pandemic disease throughout the world.
Financial support & sponsorship: Authors acknowledge financial support from the Department of Science & Technology-SERB, New Delhi and the CSIR-Institute of Genomics & Integrative Biology, Delhi, under the research project number GAP0145.
Conflicts of Interest: None.
References
- Recent advances in the development of anti-tuberculosis drugs acting on multidrug-resistant strains: A review. Int J Res Pharm Biosci. 2015;2:1-18.
- [Google Scholar]
- Survival mechanisms of pathogenic Mycobacterium tuberculosis H37Rv. FEBS J. 2010;277:2416-27.
- [Google Scholar]
- Global Tuberculosis Report 2017. Available from: https://www.who.int › publications › global_report › MainText_13Nov2017
- Host-pathogen interactions in tuberculosis. In: Cardona PJ, ed. Understanding tuberculosis – Analyzing the origin of Mycobacterium tuberculosis pathogenicity. Vienna: Intech; 2012. p. :43-76.
- [Google Scholar]
- Adhesion molecules A potent surface marker of Mycobacterium play key role in host-pathogen interaction and pathogenesis. Adv Res J Biochem Biotechnol. 2015;2:41-6.
- [Google Scholar]
- Mycobacterium tuberculosis adhesins: Potential biomarkers as anti-tuberculosis therapeutic and diagnostic targets. Microbiology. 2014;160:1821-31.
- [Google Scholar]
- Surface Proteoglycans as Mediators in Bacterial Pathogens Infections. Front Microbiol. 2016;7:220.
- [Google Scholar]
- Mycobacterial lipoarabinomannan: An extraordinary lipoheteroglycan with profound physiological effects. Glycobiology. 1998;8:113-20.
- [Google Scholar]
- Biosynthesis and virulent behavior of lipids produced by Mycobacterium tuberculosis: LAM and cord factor: An overview. Biotechnol Res Int. 2011;2011:274693.
- [Google Scholar]
- Critical roles for lipomannan and lipoarabinomannan in cell wall integrity of mycobacteria and pathogenesis of tuberculosis. MBio. 2013;4:e00472-12.
- [Google Scholar]
- Interplay between mycobacteria and host signalling pathways. Nat Rev Microbiol. 2004;2:189-202.
- [Google Scholar]
- Expression and characterization of Rv0447c product, potentially the methyltransferase involved in tuberculostearic acid biosynthesis in Mycobacterium tuberculosis. Biotechnol Appl Biochem. 2013;60:412-6.
- [Google Scholar]
- Biochemical characterization of an S-adenosyl-l-methionine-dependent methyltransferase (Rv0469) of Mycobacterium tuberculosis. Biol Chem. 2013;394:871-7.
- [Google Scholar]
- Substrate-directed function of calmodulin in autophosphorylation of Ca2+/calmodulin-dependent protein kinase II. J Biol Chem. 1998;273:28424-9.
- [Google Scholar]
- Mycobacterium tuberculosis phagosomes exhibit altered calmodulin-dependent signal transduction: Contribution to inhibition of phagosome-lysosome fusion and intracellular survival in human macrophages. J Immunol. 2001;166:3392-401.
- [Google Scholar]
- Mycobacterial lipoarabinomannans: Modulators of dendritic cell function and the apoptotic response. Microbes Infect. 2002;4:945-53.
- [Google Scholar]
- Activation of dendritic cells by toll-like receptors and C-type lectins. Handb Exp Pharmacol. 2009;188:3-30.
- [Google Scholar]
- Fibronectin: A multidomain host adhesin targeted by bacterial fibronectin-binding proteins. FEMS Microbiol Rev. 2011;35:147-200.
- [Google Scholar]
- Genetic and immunological analysis of Mycobacterium tuberculosis fibronectin-binding proteins. Infect Immun. 1991;59:2712-8.
- [Google Scholar]
- Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature. 1998;393:537-44.
- [Google Scholar]
- An overview to understand the role of PE_PGRS family proteins in Mycobacterium tuberculosis H37 Rv and their potential as new drug targets. Biotechnol Appl Biochem. 2015;62:145-53.
- [Google Scholar]
- Cloning and characterization of a novel PE_PGRS60 protein (Rv3652) of Mycobacterium tuberculosis H37 Rv exhibit fibronectin-binding property. Biotechnol Appl Biochem. 2016;63:525-31.
- [Google Scholar]
- Imperative role of fibronectin binding proteins in cell adhesion and invasion. Adv Res J Biochem Biotechnol. 2015;2:31-40.
- [Google Scholar]
- Fibronectin binding protein and Ca2+ play an access key role to mediate pathogenesis in Mycobacterium tuberculosis: An overview. Biotechnol Appl Biochem. 2016;63:820-6.
- [Google Scholar]
- The novel fibronectin-binding motif and key residues of mycobacteria. J Biol Chem. 1998;273:2905-9.
- [Google Scholar]
- Multiple moonlighting functions of mycobacterial molecular chaperones. Tuberculosis (Edinb). 2010;90:119-24.
- [Google Scholar]
- Mycobacterium tuberculosis employs cpn60.2 as an adhesin that binds CD43 on the macrophage surface. Cell Microbiol. 2010;12:1634-47.
- [Google Scholar]
- Stress wars: The direct role of host and bacterial molecular chaperones in bacterial infection. Infect Immun. 2006;74:3693-706.
- [Google Scholar]
- CD43 is required for optimal growth inhibition of Mycobacterium tuberculosis in macrophages and in mice. J Immunol. 2005;175:1805-12.
- [Google Scholar]
- Mycobacterium tuberculosis Cpn60.2 (GroEL2) blocks macrophage apoptosis via interaction with mitochondrial mortalin. Biol Open. 2017;6:481-8.
- [Google Scholar]
- Mycobacterial heat-shock protein 65 induces proinflammatory cytokines but does not activate human mononuclear phagocytes. Scand J Immunol. 1994;39:613-7.
- [Google Scholar]
- Towards understanding the biological function of the unusual chaperonin cpn60.1 (GroEL1) of Mycobacterium tuberculosis. Tuberculosis (Edinb). 2016;97:137-46.
- [Google Scholar]
- Characterization of the heparin-binding site of the mycobacterial heparin-binding hemagglutinin adhesin. J Biol Chem. 2000;275:14273-80.
- [Google Scholar]
- Identification of a heparin-binding hemagglutinin present in mycobacteria. J Exp Med. 1996;184:993-1001.
- [Google Scholar]
- Molecular characterization of the mycobacterial heparin-binding hemagglutinin, a mycobacterial adhesin. Proc Natl Acad Sci U S A. 1998;95:12625-30.
- [Google Scholar]
- Triggered Mycobacterium tuberculosis heparin-binding hemagglutinin adhesin folding and dimerization. J Bacteriol. 2011;193:2089-96.
- [Google Scholar]
- Methylation-dependent T cell immunity to Mycobacterium tuberculosis heparin-binding hemagglutinin. Nat Med. 2004;10:935-41.
- [Google Scholar]
- The heparin-binding haemagglutinin of M. tuberculosis is required for extrapulmonary dissemination. Nature. 2001;412:190-4.
- [Google Scholar]
- Characterization of fibronectin-binding antigens released by Mycobacterium tuberculosis and Mycobacterium bovis BCG. Infect Immun. 1988;56:3046-51.
- [Google Scholar]
- Evidence for three separate genes encoding the proteins of the mycobacterial antigen 85 complex. Infect Immun. 1990;58:272-4.
- [Google Scholar]
- Role of the major antigen of Mycobacterium tuberculosis in cell wall biogenesis. Science. 1997;276:1420-2.
- [Google Scholar]
- Cloning, sequence determination, and expression of a 32-kilodalton-protein gene of Mycobacterium tuberculosis. Infect Immun. 1989;57:3123-30.
- [Google Scholar]
- Disruption of the genes encoding antigen 85A and antigen 85B of Mycobacterium tuberculosis H37 Rv: Effect on growth in culture and in macrophages. Infect Immun. 2000;68:767-78.
- [Google Scholar]
- Mycolyltransferase-mediated glycolipid exchange in Mycobacteria. J Biol Chem. 2008;283:28835-41.
- [Google Scholar]
- The M. tuberculosis antigen 85 complex and mycolyltransferase activity. Lett Appl Microbiol. 2002;34:233-7.
- [Google Scholar]
- T cell response to purified filtrate antigen 85 from Mycobacterium bovis bacilli calmette-guérin (BCG) in leprosy patients. Clin Exp Immunol. 1991;86:286-90.
- [Google Scholar]
- Induction of the antigen 85 complex of Mycobacterium tuberculosis in sputum: A determinant of outcome in pulmonary tuberculosis treatment. J Infect Dis. 1998;178:1115-21.
- [Google Scholar]
- The 19-kD antigen and protective immunity in a murine model of tuberculosis. Clin Exp Immunol. 2000;120:274-9.
- [Google Scholar]
- The 19-kDa antigen of Mycobacterium tuberculosis is a major adhesin that binds the mannose receptor of THP-1 monocytic cells and promotes phagocytosis of mycobacteria. Microb Pathog. 2005;39:97-107.
- [Google Scholar]
- Mycobacterial signaling through toll-like receptors. Front Cell Infect Microbiol. 2012;2:145.
- [Google Scholar]
- Mycobacterium tuberculosis 19-kDa lipoprotein promotes neutrophil activation. J Immunol. 2001;167:1542-9.
- [Google Scholar]
- Toll-like receptor 2-dependent inhibition of macrophage class II MHC expression and antigen processing by 19-kDa lipoprotein of Mycobacterium tuberculosis. J Immunol. 2001;167:910-8.
- [Google Scholar]
- The Mycobacterium tuberculosis 19-kilodalton lipoprotein inhibits gamma interferon-regulated HLA-DR and fc gamma R1 on human macrophages through toll-like receptor 2. Infect Immun. 2003;71:4487-97.
- [Google Scholar]
- Mycobacterial 19-kDa lipoprotein mediates Mycobacterium tuberculosis-induced apoptosis in monocytes/macrophages at early stages of infection. Cell Death Differ. 2000;7:1270-2.
- [Google Scholar]
- A systems chemical biology study of malate synthase and isocitrate lyase inhibition in Mycobacterium tuberculosis during active and NRP growth. Comput Biol Chem. 2013;47:167-80.
- [Google Scholar]
- Mycobacterium tuberculosis malate synthase is a laminin-binding adhesin. Mol Microbiol. 2006;60:999-1013.
- [Google Scholar]
- Role of glyoxylate shunt in oxidative stress response. J Biol Chem. 2016;291:11928-38.
- [Google Scholar]
- Glyoxylate metabolism and adaptation of Mycobacterium tuberculosis to survival under anaerobic conditions. Infect Immun. 1982;37:1042-9.
- [Google Scholar]
- Mycobacterium tuberculosis malate synthase- and MPT51-based serodiagnostic assay as an adjunct to rapid identification of pulmonary tuberculosis. Clin Vaccine Immunol. 2006;13:1291-3.
- [Google Scholar]
- Mycobacterium tuberculosis produces pili during human infection. Proc Natl Acad Sci U S A. 2007;104:5145-50.
- [Google Scholar]
- Mycobacterium tuberculosis pili promote adhesion to and invasion of THP-1 macrophages. Jpn J Infect Dis. 2014;67:476-8.
- [Google Scholar]
- Evaluation of the role of Mycobacterium tuberculosis pili (MTP) as an adhesin, invasin, and cytokine inducer of epithelial cells. Braz J Infect Dis. 2016;20:160-5.
- [Google Scholar]
- Glyceraldehyde-3-phosphate dehydrogenase of Paracoccidioides brasiliensis is a cell surface protein involved in fungal adhesion to extracellular matrix proteins and interaction with cells. Infect Immun. 2006;74:382-9.
- [Google Scholar]
- Mycobacterium tuberculosis acquires iron by cell-surface sequestration and internalization of human holo-transferrin. Nat Commun. 2014;5:4730.
- [Google Scholar]
- Role of extracellular GAPDH in Streptococcus pyogenes virulence. Mo Med. 2013;110:236-40.
- [Google Scholar]
- Kinetic and mechanistic characterization of the glyceraldehyde 3-phosphate dehydrogenase from Mycobacterium tuberculosis. Arch Biochem Biophys. 2013;540:53-61.
- [Google Scholar]
- Role of apoptosis signal-regulating kinase 1 (ASK1) as an activator of the GAPDH-siah1 stress-signaling cascade. J Biol Chem. 2015;290:56-64.
- [Google Scholar]
- Targeting iron acquisition by Mycobacterium tuberculosis. Infect Disord Drug Targets. 2007;7:213-20.
- [Google Scholar]
- Over-expression of GAPDH induces apoptosis in COS-7 cells transfected with cloned GAPDH cDNAs. Neuroreport. 1999;10:2029-33.
- [Google Scholar]




