
Translate this page into:
Phenotypic & genotypic study of antimicrobial profile of bacteria isolates from environmental samples
For correspondence: Dr Indrani Karunasagar, UNESCO MIRCEN for Medical & Marine Biotechnology, Center for Science Education & Research, Nitte (Deemed to be University), University Enclave, Medical Sciences Complex, Deralakatte, Mangaluru 575 018, Karnataka, India e-mail: indrani.karunasagar@nitte.edu.in
-
Received: ,
This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
This article was originally published by Wolters Kluwer - Medknow and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Background & objectives:
The resistance to antibiotics in pathogenic bacteria has increased at an alarming rate in recent years due to the indiscriminate use of antibiotics in healthcare, livestock and aquaculture. In this context, it is necessary to monitor the antibiotic resistance patterns of bacteria isolated from the environmental samples. This study was conducted to determine the phenotypic and genotypic profile of antimicrobial resistance in Gram-negative bacteria isolated from environmental samples.
Methods:
Two hundred and fifty samples were collected from different sources, viz. fish and fishery products (99), livestock wastes (81) and aquaculture systems (70), in and around Mangaluru, India. Isolation, identification and antimicrobial profiling were carried out as per standard protocols. The isolates were screened for the presence of resistance genes using PCR.
Results:
A total of 519 Gram-negative bacteria comprising Escherichia coli (116), Salmonella spp. (14), Vibrio spp. (258), Pseudomonas spp. (56), Citrobacter spp. (26) and Proteus spp. (49) were isolated and characterized from 250 samples obtained from different sources. A total of 12 antibiotics were checked for their effectiveness against the isolates. While 31.6 per cent of the isolates were sensitive to all the antibiotics used, 68.4 per cent of the isolates showed resistance to at least one of the antibiotics used. One-third of the isolates showed multidrug resistance. Maximum resistance was observed for ampicillin (43.4%), followed by nitrofurantoin (20.8%). Least resistance was seen for carbapenems and chloramphenicol. PCR profiling of the resistant isolates confirmed the presence of resistance genes corresponding to their antibiotic profile.
Interpretation & conclusions:
This study results showed high rate of occurrence of antimicrobial resistance and their determinants in Gram-negative bacteria isolated from different environmental sources.
Keywords
Antimicrobial resistance
environmental samples
genotypic characterization
multidrug resistance

The development and spread of antibiotic resistance among bacteria affecting human health are most challenging problems of the modern world. The genetic determinants responsible for conferring resistance are often carried in self-transmissible mobile elements such as conjugative plasmids, gene cassettes in integrons and transposons, which are transferred between bacterial species, resulting in transmission of resistance to other species1.
The development of resistance to a specific antimicrobial compound is influenced by several environmental factors and it has been reported that the organisms isolated from environment with high faecal contamination can easily acquire resistance to the common antimicrobial drugs2. In addition, the non-therapeutic use of antimicrobials in animal husbandry, aquaculture, agriculture, poultry and piggery has increased the incidence of multidrug resistance forms in the environment34. The presence of antibiotic resistance in Gram-negative bacteria has been well documented including their occurrence in aquaculture5 and livestock6. Among the Gram-negative bacteria, the prevalence of antimicrobial resistance has been reported to be high in Escherichia coli7, Salmonella spp.8, Vibrio spp.9, Pseudomonas spp.10, Citrobacter spp.11 and Proteus spp.12. It is important to understand and monitor the antibiotic resistance patterns of human pathogenic bacteria persisting in the environment. Hence, the main objective of this study was to investigate the phenotypic and genotypic profile of antimicrobial resistance in Gram-negative bacteria isolated from environmental samples.
Material & Methods
A total of 250 samples from different sources, viz. 99 from fish and fishery products comprising 61 of fish/shellfish and 38 of oyster/clam/molluscs; 81 from livestock wastes comprising poultry (23), piggery (14) and cattle wastes (44) samples and 70 from aquaculture systems comprising fish farm water (40) and pond sediment (30), in and around Mangaluru, India, were aseptically collected fortnightly during the study period (2011-2014). The samples were subjected to isolation of associated bacteria by culture-based conventional methods13. Typical colonies from the selective plates were sub-cultured onto Luria Bertani (L-B) agar (HiMedia Laboratories Pvt. Ltd., Mumbai) and identified using standard biochemical tests, viz. Gram staining, motility, cytochrome oxidase, catalase, oxidation fermentation test, urease and triple sugar iron agar1415. E. coli, Salmonella spp. and Vibrio spp. were further confirmed by single-step PCR using species-specific primers (Table I)161718. Identified isolates were preserved in 30 per cent glycerol L-B broth and stored at −80°C for further studies.
| Isolate | Gene | Sequence 5’- 3’ | Size (bp) | Tm (°C) | Reference |
|---|---|---|---|---|---|
| Escherichia coli | uidA | AAAACGGCAAGAAAAAGCAG | 146 | 60 | 16 |
| ACGCGTGGTTACAGTCTTGCG | |||||
| Salmonella | invA | GTGAAATTATCGCCACGTTCGGGCAA | 284 | 64 | 17 |
| TCATCGCACCGTCAAAGGAACC | |||||
| hns | TACCAAAGCTAAACGCGCAGCT | 156 | 60 | 18 | |
| TGATCAGGAAATCTTCCAGTTGC | |||||
| Vibrio | 16S rRNA | GTAAATTATCGCCACGTTCGG | 321 | 55 | 16 |
| AACGGCAAGAAAAAGCAGTG |
Tm, melting/annealing temperature of primers
Antibiotic susceptibility test: Antibiotic susceptibility tests were performed for all the isolates using the disc diffusion method described by Bauer et al19 as per the Clinical and Laboratory Standards Institute guidelines20. Twelve common antibiotics (in μg) namely nalidixic acid (30), tetracycline (30), co-trimoxazole (25), ciprofloxacin (5), chloramphenicol (30), ampicillin (10), gentamicin (10), nitrofurantoin (300), imipenem (10), meropenem (10mc), cefotaxime (30) and piperacillin-tazobactam (100/10) (HiMedia) were used for antibiotic profiling. ATCC 25922 E. coli culture was used as a standard quality control strain for AST for Enterobacteriaceae groups.
Detection of antibiotic resistance determinants by PCR: The isolates showing resistance to particular antibiotics were selected and screened for the presence of antibiotic resistance determinants by PCR using the primers listed in Table II21222324. Genomic DNA was extracted from the bacterial culture by cetyl-trimethyl ammonium bromide method25. Using genomic DNA as a template, PCR was carried out in 30 μl reaction mixture containing 10× buffer (100 mM Tris-HCl, p H 8.3, 20 mM MgCl2, 500 mM KCl, 0.1% gelatin), 200 mM of dNTPs (dATP, dTTP, dGTP and dCTP), 10 pmol each of forward and reverse primers and 1.0 unit of Taq DNA polymerase enzyme (HiMedia) in a MJ-Research Thermo Cycler (PTC-200, Bio-Rad, USA). The PCR conditions included initial denaturation at 94°C for 5 min, followed by 35 cycles with each cycle consisting of denaturation at 94°C for 60 sec, annealing for 60 sec at an optimized temperature depending on the primer set used and extension at 72°C for 30 sec. The final extension was set at 72°C for 10 min.
| Antimicrobials | Resistance genes | Forward and reverse primer 5’- 3’ | Size (bp) | Present in samples | Reference |
|---|---|---|---|---|---|
| Tetracycline | tetA | TTGGCATTCTGCATTCACTC | 494 | Yes | 21 |
| GTATAGCTTGCCGGAAGTCG | |||||
| tetB | CAGTGCTGTTGTTGTCATTAA | 571 | Yes | ||
| GCTTGGAATACTGAGTGTTAA | |||||
| tetC | CTTGAGAGCCTTCAACCCAG | 418 | Yes | ||
| ATGGTCGTCATCTACCTGCC | |||||
| tetD | GCAAACCATTACGGCATTCT | 546 | Yes | ||
| GATAAGCTGCGCGGTAAAAA | |||||
| tetE | TATTAACGGGCTGGCATTTC | 544 | Yes | ||
| AGCTGTCAGGTGGGTCAAAC | |||||
| tetG | GCTCGGTGGTATCTCTGCTC | 550 | Yes | ||
| CAAAGCCCCTTGCTTGTTAC | |||||
| tetL | CATTTGGTCTTATTGGATCG | 488 | No | 22 | |
| ATTACACTTCCGATTTCGG | |||||
| tetM | GTTAAATAGTGTTCTTGGAG | 657 | Yes | ||
| CTAAGATATGGCTCTAACAA | |||||
| tetS | TGGAACGCCAGAGAGGTATT | 660 | Yes | ||
| ACATAGACAAGCCGTTGACC | |||||
| Sulphonamides | sul I | TTTCCTGACCCTGCGCTCTAT | 425 | Yes | 21 |
| GTGCGGACGTAGTCAGCGCCA | |||||
| sul II | CCTGTTTCGTCCGACACAGA | 435 | Yes | ||
| GAAGCGCAGCCGCAATTCAT | |||||
| sul III | ATGAGCAAGATTTTTGGAATCGTAA | 792 | Yes | ||
| CTAACCTAGGGCTTTGGATATTT | |||||
| Chloramphenicol | cat1 | AACCAGACCGTTCAGCTGGAT | 549 | Yes | |
| CCTGCCACTCATCGCAGTAC | |||||
| cat2 | AACGGCATGATGAACCTGAA | 547 | Yes | ||
| ATCCCAATGGCATCGTAAAG | |||||
| cat3 | ATCGGCATCGGTTACCATGT | 531 | No | ||
| ATCCCCTTCTTGCTGATATT | |||||
| cmlA | GGCCTCGCTCTTACGTCATC | 662 | Yes | ||
| GCGACACCAATACCCACTAGC | |||||
| cmlB | ACTCGGCATGGACATGTACT | 840 | No | ||
| ACGGACTGCGGAATCCATAG | |||||
| floR | ATGACCACCACACGCCCCG | 1,213 | No | ||
| AGACGACTGGCGACTTCTCG | |||||
| Quinolones | qnrA | ATTTCTCACGCCAGGATTTG | 516 | Yes | 23 |
| GATCGGCAAAGGTTAGGTCA | |||||
| qnrB | GATCGTGAAAGCCAGAAAGG | 469 | Yes | ||
| ACGATGCCTGGTAGTTGTCC | |||||
| qnrS | ACGACATTCGTCAACTGCAA | 417 | Yes | ||
| TAAATTGGCACCCTGTAGGC | |||||
| Ampicillin | blaTEM | CTCACCCAGAAACGCTGGTG | 569 | Yes | 24 |
| ATCCGCCTCCATCCAGTCTA | |||||
| Cefotaxime | blaCTX-M | ACGTTAAACACCGCCATTCC | 356 | Yes | |
| TCGGTGACGATTTTAGCCGC |
Sequencing of antibiotic-resistant determinants: The amplified PCR products (antibiotic-resistant determinants) were purified using PCR Purification Kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol and were outsourced for capillary sequencing. The obtained sequences were analyzed by BLAST (http://blast.ncbi.nlm.nih.gov) and antibiotic resistance genes database for their homology with the database sequences and the confirmed sequences were submitted to the GenBank.
Results
Five hundred and nineteen bacterial strains were isolated and identified to the genus level by conventional methods using a battery of biochemical tests. The details of the isolates identified from the different sources are given in Table III. Molecular confirmation of E. coli, Vibrio spp. and Salmonella spp. was done by PCR using primers namely uidA for E. coli, invA and hns for Salmonella and 16S rRNA for Vibrio (Fig. 1).
| Source | Number of samples | Escherichia coli | Vibrio spp. | Salmonella spp. | Pseudomonas spp. | Citrobacter spp. | Proteus spp. | Total |
|---|---|---|---|---|---|---|---|---|
| Fish and fishery products (fish/shrimps and clams/oysters/mussels) | 99 | 45 | 169 | 8 | 18 | 6 | 13 | 259 |
| Livestock wastes (poultry, piggery and cattle farm) | 81 | 61 | 7 | 6 | 17 | 10 | 18 | 119 |
| Environmental samples (farm water and pond sediment) | 70 | 10 | 82 | - | 21 | 10 | 18 | 141 |
| Total | 250 | 116 | 258 | 14 | 56 | 26 | 49 | 519 |
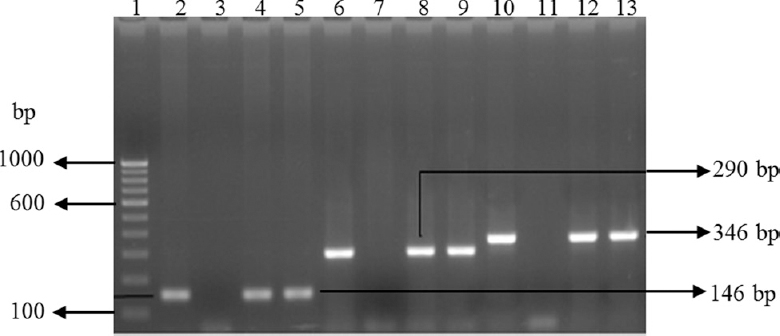
- Gel-electrophoresis of PCR amplified products of Escherichia coli and Salmonella isolates. Lane1: 100 bp DNA ladder; lane 2: uidA gene-positive control (E. coli); lane 3: uidA gene-negative control (E. coli); lanes 4 & 5: uidA gene-positive E. coli isolates; lane 6: positive control for invA gene (Salmonella); lane 7: negative control for invA; lanes 8 & 9: invA gene-positive Salmonella isolates; lane 10: positive control for hns gene (Salmonella); lane 11: negative control for hns gene; lanes 12-13: hns gene-positive Salmonella spp. isolates.
Antimicrobial susceptibility test: Among the 519 isolates tested for antimicrobial susceptibility, only 31.6 per cent (164 isolates) showed susceptibility to all the antibiotics tested. While 68.4 per cent (355 isolates) were resistant to at least one of the antibiotics, 33.7 per cent (175 isolates) displayed multidrug resistance (resistance to more than one antibiotic). As shown in Fig. 2, maximum resistance was observed for ampicillin (225 isolates, 43.4%) followed by nitrofurantoin (108 isolates, 20.8%), nalidixic acid (71 isolates, 13.7%), cefotaxime (62 isolates, 11.9%), tetracycline (58 isolates, 11.2%), co-trimoxazole (38 isolates, 7.3%), ciprofloxacin (30 isolates, 5.8%), gentamicin and piperacillin/tazobactam (28 isolates, 5.4%), imipenem (18 isolates, 3.5%), meropenem (16 isolates, 3.1%) and chloramphenicol (15 isolates, 2.9%).
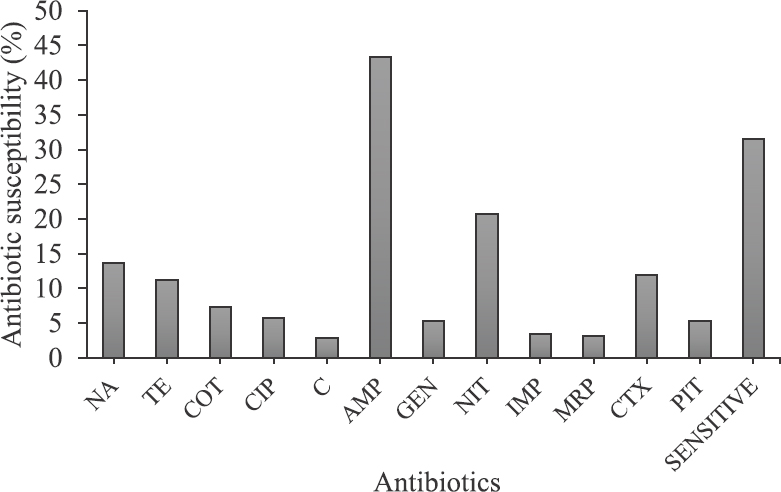
- Antibiotic resistance pattern of all isolates against 12 different antibiotics used in the study. NA, nalidixic acid; TE, tetracycline; COT, co-trimoxazole; CIP, ciprofloxacin; C, chloramphenicol; AMP, ampicillin; GEN, gentamicin; NIT, nitrofurantoin; MRP, meropenem; CTX, cefotaxime; PIT, piperacillin/tazobactum.
The drug resistance patterns of the isolates according to the source of collection are presented in Fig. 3. Among isolates obtained from fishery products and environment samples, 47.5 per cent showed resistance to ampicillin and 18 per cent to nitrofurantoin, suggesting maximum resistance to these two antibiotics. Among isolates from livestock wastes, maximum resistance was observed for nitrofurantoin (33.6% of the isolates) followed by ampicillin (28.6% of the isolates) and tetracycline (22.7% of the isolates).
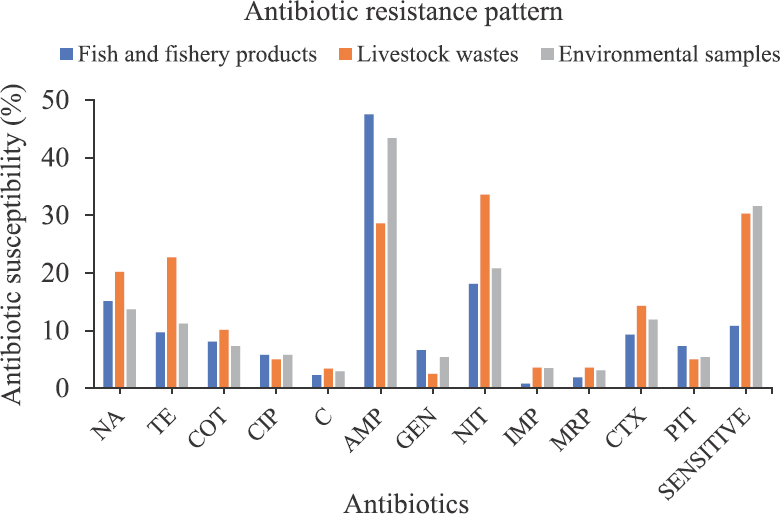
- Source-wise representation of antibiotic resistance pattern of the isolates to 12 antibiotics. Abbreviations are as given in Fig. 2.
Detection of antibiotic resistance genes: The antibiotic-resistant determinants associated with resistance were detected by PCR. Of the 58 isolates showing resistance to tetracycline, 45 (77.6%) harboured one or more than one tetracycline-resistant genes (tetA, tetB, tetC, tetD, tetE, tetG, tetM and tetS). Remaining 13 isolates did not carry any of the tet genes tested even though these were phenotypically resistant. Among 38 isolates resistant to co-trimoxazole, 12 (31.6%) harboured at least one of the sul genes (sul1, sul2, sul3) and the 26 isolates did not harbour any of the tested genes (Fig. 4). Two hundred and twenty five ampicillin-resistant isolates were tested for the presence of blaTEM, the gene responsible for the resistance. However, only eight isolates (3.6%) showed the presence of this gene. Of the 62 cefotaxime-resistant isolates, only five (8%) had the blaCTX-M gene conferring the resistance trait. Among the 15 isolates resistant to chloramphenicol, five (33.3%) carried one of the resistance genes (cat1, cat2 and cmlA), but none of these showed the presence of cat3, cmlB and floR (Fig. 5). Of the 71 nalidixic acid-resistant isolates, nine (12.7%) carried either qnrA, qnrB or qnrS.

- Detection of PCR amplified products of antibiotic resistance genes. Lane 1: 100 bp DNA ladder; lane 2: tetA (492 bp); lane 4: tetB (571 bp); lane 6: tetE (544 bp); lane 8: sul1 (425 bp); lane 10: sul2 (435 bp); lane 12: sul3 (792 bp); lane 14: 500 bp DNA ladder and lanes 3, 5, 7, 9, 11, 13: negative controls.
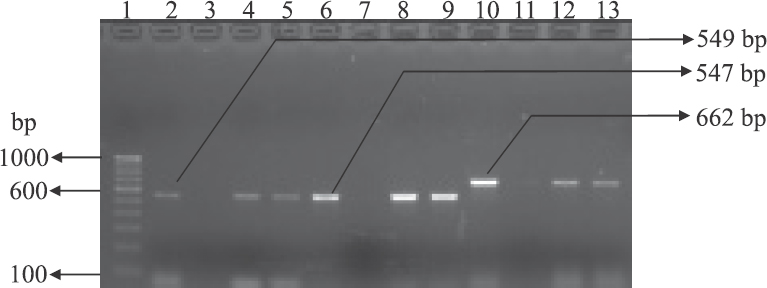
- Gel-electrophoresis of PCR amplified products of chloramphenicol-resistant isolates. Lane 1: 100 bp DNA ladder; lane 2: cat1 gene-positive control; lane 3: cat1 gene-negative control; lanes 4-5: cat1 gene-positive isolates; lane 6: positive control for cat2 gene; lane 7: negative control for cat2 gene; lanes 8-9: cat2 gene-positive isolates; lane 10: positive control for cmlA gene; lane 11: negative control for cmlA gene; lane 12-13: cmlA gene-positive isolates.
Further, sequencing of the PCR products of antibiotic resistance genes from representative isolates revealed 98 per cent identity with the existing antibiotic resistance gene sequences in the database. The GenBank accession numbers of these sequences are given in Table IV.
| Title | Accession number |
|---|---|
| Escherichia coli isolate T1 tetracycline resistance protein class A (tetA) gene, partial cds. (EC12) | KF240812.1 |
| E. coli strain T2 tetracycline resistance protein class B (tetB) gene, partial cds. (EC23) | KF240811.1 |
| Salmonella sp. T1a tetracycline resistance proteins class A (tetA) gene, partial cds. (S131) | KF240813.1 |
| Salmonella sp. S1a sulphonamide resistance protein (sul1) gene, partial cds. (S131) | KF240817.1 |
| E. coli strain S2 sulphonamide resistance protein (sul2) gene, partial cds. (EC11) | KF240815.1 |
| E. coli strain S3 sulphonamide resistance protein (sul3) gene, partial cds. (EC10) | KF240814.1 |
| E. coli strain S1 sulphonamide resistance protein (sul1) gene, partial cds. (EC10) | KF240816.1 |
Discussion
The results of antibiotic susceptibility test revealed that the number of isolates showing resistance to one or more antibiotics was on the rise, suggesting the high occurrence of antibiotic resistance in animal, wastewater, soil and other natural environments. Regardless of their source of isolation, resistance to three antibiotics, namely ampicillin, nitrofurantoin and tetracycline, was most frequently observed. Although ampicillin resistance was highest among all the antibiotics used.
It has been shown that isolates from fish and shrimp farms have widespread resistance to nitrofurantoin26. In this study, approximately 20 per cent of the isolates showed resistance to nitrofurantoin. Similarly, 18 per cent of the isolates obtained from fish farms and related products showed resistance to this antibiotic, and the results were in agreement with that of an earlier study27. The resistance to nitrofurantoin could be either due to the inhibition of nitrofurantoin reductase or due to the nucleotide changes (mutation) in nfsA and nfsB encoding oxygen-insensitive nitro reductase. Nitrofurantoin and its metabolites have zero tolerance in fishery products, and yet the high occurrence of resistance to nitrofurantoin observed in the study indicating the use of this antibiotic in aquaculture and other environments.
Resistance to tetracycline has been reported frequently from environmental samples28. In the present study, 11.2 per cent of the isolates exhibited phenotypic resistance to tetracycline, which was consistent with earlier studies that reported tetracycline resistance in 15-56 per cent of the isolates28. Resistance to tetracycline is more frequently observed in poultry isolates because it is widely used as growth promoters in poultry production29. More than 60 per cent of poultry isolates showed resistance to tetracycline in this study.
Skockova et al30 had earlier reported that tetA and tetB were the most common genes responsible for resistance to tetracycline. In our study, 77 per cent of the phenotypically resistant isolates showed the presence of one or more tet genes. Although the remaining 23 per cent did not carry any of the tet genes tested in the study despite being phenotypically resistant, the resistance in such isolates could be due to other mechanisms such as enzymatic inactivation or target modification31.
In this study, 7.3 per cent of the total isolates showed moderate resistance to co-trimoxazole. The sulphonamide resistance genes (sul1, sul2 and sul3) are known to be associated with class 1 integrons responsible for capturing and excision of genes during site-specific recombination events32, which further facilitates the emergence of multidrug resistance among bacterial pathogens. Similar to the pattern seen for other antibiotics, the phenotypic and genotypic association was inconsistent for co-trimoxazole with only 31.6 per cent of the phenotypically confirmed isolates showing the presence of one of the three sul genes. The absence of sul in the remaining isolates suggests the possibility of some new genes or resistance mechanism to confer resistance to co-trimoxazole.
The antibiotic resistance pattern obtained for chloramphenicol in this study was low profile i.e.,2.9 per cent. The results of genotypic characterization showed inconsistency with 20 per cent carrying both type 1 and type 2 cat genes and 13.3 per cent harbouring cmlA. However, none of the isolates were positive for type 3 cat, cmlB and floR. The absence of resistance genes was observed in 33.3 per cent of the phenotypically resistant isolates, suggesting the possibility of other mechanism(s) such as overexpression of efflux pumps, mutations or modifications in the target sites or decreased outer membrane permeability contributing to chloramphenicol resistance3334.
The resistance to β-lactam group of antibiotics such as ampicillin, cefotaxime, imipenem, meropenem and piperacillin/tazobactam was also analyzed in this study. Except for ampicillin, which showed maximum resistance, the percentage of resistance was relatively less with 3.5 and 3.1 per cent of isolates showing resistance to imipenem and meropenem, respectively. In our study, 20 per cent of the isolates showed resistance to nalidixic acid and five per cent towards ciprofloxacin. The usual cause of resistance to quinolones is due to point mutation/mutations in the quinolone resistance determining regions (QRDR) or due to presence of active efflux or outer membrane permeability. In addition, the plasmid-mediated quinolone resistance (PMQR) has also been reported. In the present study, nalidixic acid- and ciprofloxacin-resistant isolates carried qnrA, qnrB and qnrS indicating that quinolone resistance was acquired through plasmid-mediated determinant. Although point mutations in the QRDR are the main reason for resistance to quinolone/fluoroquinolones, the occurrence of PMQR genes cannot be neglected since these play a major role in the transmission of resistance among bacterial isolates35. The bacterial isolates showed similar pattern of resistance to cell wall synthesis inhibitors such as meropenem and imipenem. Although the susceptibility of the majority of the isolates to carbapenems was encouraging, the small percentage of resistance observed should be viewed seriously.
In conclusion, the results of this study on the pattern of resistance to antimicrobials in environmental samples highlighted the importance of continuous vigilance on the distribution of multidrug-resistant human pathogens in the environment. High rate of multidrug resistance among bacterial isolates from environmental samples suggested the indiscriminate use of antimicrobials in various sectors of animal husbandry. Judicious use of antibiotics, use of alternative bio-control approaches or development of pathogen-specific antimicrobial agents could help in combating antimicrobial resistance.
Financial support & sponsorship: Financial support received by the last author (IK) from the Indian Council of Medical Research (Grant no. AMR/37/2011-ECD-I), New Delhi, towards this study and the research fellowship to the first author (SKS) is acknowledged.
Conflicts of Interest: None.
References
- Prevalence of antibiotic resistance in drinking water treatment and distribution systems. Appl Environ Microbiol. 2009;75:5714-8.
- [Google Scholar]
- The routine use of antibiotics to promote animal growth does little to benefit protein undernutrition in the developing world. Clin Infect Dis. 2005;41:1007-13.
- [Google Scholar]
- Antibiotic growth promoters in agriculture: History and mode of action. Poult Sci. 2005;84:634-43.
- [Google Scholar]
- Antibiotic resistance in Gram-negative bacteria isolated from farmed catfish. Food Control. 2007;18:1391-6.
- [Google Scholar]
- Phenotypic and genotypic antimicrobial resistance and virulence genes of Salmonella enterica isolated from pet dogs and cats. J Vet Sci. 2017;18:273-81.
- [Google Scholar]
- Investigation of stx 2+ eae+ Escherichiacoli O157: H7 in beef imported from Malaysia to Thailand. Int Food Res J. 2011;18:381-6.
- [Google Scholar]
- Characterization of multiple-antimicrobial-resistant Salmonella serovars isolated from retail meats. Appl Environ Microbiol. 2004;70:1-7.
- [Google Scholar]
- Antibiotic resistance of Vibrio cholerae from poultry sources of Dhaka, Bangladesh. Adv Biol Res. 2008;2:60-7.
- [Google Scholar]
- Antibiotic resistance of Pseudomonas aeruginosa in pneumonia at a Single University Hospital Center in Germany over a 10-year period. PLoS One. 2015;10:e0139836.
- [Google Scholar]
- The antibiotic resistance pattern in Citrobacter species: An emerging nosocomial pathogen in a tertiary care hospital. J Clin Diagn Res. 2012;6:642-4.
- [Google Scholar]
- Occurrence, species distribution and antibiotic resistance of Proteus isolates: A case study at the Komfo Anokye Teaching Hospital (KATH) in Ghana. Int J Pharm Sci Res. 2010;1:347-52.
- [Google Scholar]
- Salmonella. In: Bacteriological analytical manual. Gaithersburg, MD: Federal Drug Administration; 2011.
- [Google Scholar]
- Basic microbiology: A illustrated laboratory manual. New Delhi, India: Daya Publishing House; 2011.
- [Google Scholar]
- Biochemical tests for identification of medical bacteria. Philadelphia: Williams & Wilkins Co; 1976.
- [Google Scholar]
- Detection of Escherichiacoli and Shigella spp. in water by using the polymerase chain reaction and gene probes for uid. Appl Environ Microbiol. 1991;57:1013-7.
- [Google Scholar]
- Amplification of an InvA gene sequence of Salmonella typhimurium by polymerase chain reaction as a specific method of detection of Salmonella. Mol Cell Probes. 1992;6:271-9.
- [Google Scholar]
- Detection of Salmonella spp. in oysters using polymerase chain reaction (PCR) and gene probes. J Food Sci. 1993;58:1191-7.
- [Google Scholar]
- Antibiotic susceptibility testing by a standardized single disk method. Am J Clin Pathol. 1966;45:493-6.
- [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance standards for antimicrobial disk susceptibility tests. In: CLSI Document M100 (28thed). Wayne, PA: CLSI; 2018.
- [Google Scholar]
- Detection of antimicrobial resistance genes of pathogenic Salmonella from swine with DNA microarray. J Vet Diagn Invest. 2007;19:161-7.
- [Google Scholar]
- Comparison of antimicrobial resistance phenotypes and resistance genes in Enterococcus faecalis and Enterococcus faecium from humans in the community, broilers, and pigs in Denmark. Diagn Microbiol Infect Dis. 2000;37:127-37.
- [Google Scholar]
- Qnr prevalence in ceftazidime-resistant Enterobacteriaceae isolates from the United States. Antimicrob Agents Chemother. 2006;50:2872-4.
- [Google Scholar]
- Antibiotic resistance genes in the bacteriophage DNA fraction of environmental samples. PLoS One. 2011;6:e17549.
- [Google Scholar]
- Current protocols in molecular biology. (4th ed). New Jersey: John Wiley & Sons; 1995.
- [Google Scholar]
- Plasmid-mediated antibiotic resistance in Pseudomonas (Burkholderia) cepacia from brackish water aquaculture farms. Indian J Fish. 2006;53:263-9.
- [Google Scholar]
- High resistance prevalence towards ampicillin, co-trimoxazole and ciprofloxacin, among uropathogenic Escherichia coli isolates in Mexico city. J Infect Dev Ctries. 2008;2:350-3.
- [Google Scholar]
- Multiple antimicrobial resistance in Vibrio spp. isolated from river and aquaculture water sources in Imo state, Nigeria. J Br Microbiol Res. 2014;4:560-9.
- [Google Scholar]
- Occurrence of antibiotic-resistant bacterial strains isolated in poultry. Vet Med Czech. 2002;47:52-9.
- [Google Scholar]
- Detection of tetracycline resistance genes in Escherichia coli from raw cow's milk. J Microbiol Biotechnol Food Sci. 2012;1:777.
- [Google Scholar]
- Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol Mol Biol Rev. 2001;65:232-60.
- [Google Scholar]
- Detection of class 1 integrons in Salmonella weltevreden and silent antibiotic resistance genes in some seafood-associated nontyphoidal isolates of salmonella in South-West coast of India. J Appl Microbiol. 2012;112:1113-22.
- [Google Scholar]
- Adaptive drug resistance mediated by root-nodulation-cell division efflux pumps. Clin Microbiol Infect. 2009;15(Suppl 1):32-6.
- [Google Scholar]
- Responses of wild-type and resistant strains of the hyperthermophilic bacterium Thermotoga maritima to chloramphenicol challenge. Appl Environ Microbiol. 2007;73:5058-65.
- [Google Scholar]
- Multiple antimicrobial resistance and novel point mutation in fluoroquinolone-resistant Escherichiacoli isolates from Mangalore, India. Microb Drug Resist. 2017;23:994-1001.
- [Google Scholar]




