
Translate this page into:
PI3K/Akt/mTOR signaling & its regulator tumour suppressor genes PTEN & LKB1 in human uterine leiomyomas
Reprint requests: Dr Madhu Mati Goel, Department of Pathology, King George's Medical University, Lucknow 226 003, Uttar Pradesh, India e-mail: madhukgmc@gmail.com
-
Received: ,
This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under the identical terms.
This article was originally published by Medknow Publications & Media Pvt Ltd and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Background & objectives:
Despite their high occurrence and associated significant level of morbidity manifesting as spectrum of clinical symptoms, the pathogenesis of uterine leiomyomas (ULs) remains unclear. We investigated expression profile of tumour suppressor genes PTEN (phosphatase and tensin homolog deleted on chromosome ten) and LKB1 (liver kinase B1), and key signaling components of P13K (phosphatidylinositol 3-kinase)/Akt (protein kinase B)/mTOR (mammalian target of rapamycin) pathway in leiomyomas and adjacent normal myometrium in women of reproductive age, to explore the possibility of targeting this pathway for future therapeutic implications.
Methods:
Real time PCR (qPCR) was used to quantify relative gene expression levels of PTEN, Akt1, Akt2, mTOR, LKB1 and VEGFA (vascular endothelial growth factor A) in leiomyoma as compared to adjacent normal myometrium. Immunohistochemistry was subsequently performed to analyze expression of PTEN, phospho-Akt, phospho-mTOR, phospho-S6, LKB1 and VEGFA in leiomyoma and adjacent normal myometrium.
Results:
Significant upregulation of PTEN (2.52 fold; P=0.03) and LKB1 (3.93 fold; P=0.01), and downregulation of VEGFA (2.95 fold; P=0.01) genes were observed in leiomyoma as compared to normal myometrium. Transcript levels of Akt1, Akt2 and mTOR did not vary significantly between leiomyoma and myometrium. An increased immunoexpression of PTEN (P=0.015) and LKB1 (P<0.001) and decreased expression of VEGFA (P=0.01) was observed in leiomyoma as compared to myometrium. Immunostaining for activated (phosphorylated) Akt, mTOR and S6 was absent or low in majority of leiomyoma and myometrium.
Interpretation & conclusions:
Upregulation of PTEN and LKB1 in concert with negative or low levels of activated Akt, mTOR and S6 indicates that PI3K/Akt/mTOR pathway may not play a significant role in pathogenesis of leiomyoma.
Keywords
Immunohistochemistry
patient matched myometrium
PI3K/Akt/mTOR signaling pathway
quantitative real time PCR
tumour suppressor genes
uterine leiomyomas

Uterine leiomyomas, also known as myomas or fibroids are the most common pelvic tumours occurring in up to 77 per cent of women and are the leading indication for hysterectomy worldwide. Though benign, these significantly affect the reproductive health of women, due to their high incidence and the lack of proven non-surgical treatment options. Leiomyomas are often associated with a variety of clinical symptoms including excessive menstrual bleeding, dysmenorrhoea, chronic pelvic pain and pressure symptoms leading to morbidity1.
Despite their high prevalence, the pathogenesis of uterine leiomyoma remains unclear. Recent progresses in defining the molecular mechanisms underlying the growth and development of leiomyomas have suggested that leiomyomas grow under the control of ovarian hormones which act by regulating the growth factors and their signaling pathways2. Elevated levels of both oestrogen and progesterone receptors have been demonstrated in leiomyoma as compared to autologous myometrium suggesting that leiomyomas are capable of a heightened response to these hormones3. Studies in rat uterine cells and breast cancer cells have shown that oestrogen and progesterone can rapidly activate the phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathway45. Dysregulation of PI3K/Akt pathway has been implicated in many cancers such as breast, endometrium, prostate and ovary6 and more recently in benign proliferative disorders such as lymphangioleiomyomatosis, tuberous sclerosis complex syndrome and uterine leiomyomata7. Differential expressions of PI3K/Akt pathway proteins in patient-matched leiomyoma and myometrial tissues have been reported8.
Activated PI3K catalyzes the production of phosphatidylinositol-3,4,5-triphosphate (PIP3) at the cell membrane, which in turn leads to recruitment, phosphorylation, and activation of the serine/threonine protein kinase B (Akt). The tumour suppressor gene PTEN (phosphatase and tensin homolog deleted on chromosome ten) negatively regulates phosphorylation of Akt. Inactivating mutations of PTEN or constitutive activation of growth factor receptors and PI3K result in increased requirement of Akt phosphorylation for cell growth in different cancer and cancer predisposition syndromes9. Once activated, signaling through Akt can be propagated to varied substrates including mTOR (mammalian target of rapamycin), a key regulator of protein translation at least in part, through inhibition of tuberous sclerosis complex-2 (TSC2) gene product tuberin. TSC2 is one of two tumour suppressor genes found mutated in the tuberous sclerosis complex disease8. TSC2 is also regulated by adenosine monophosphate-activated protein kinase (AMPK), which phosphorylates TSC2 to activate its tumour suppressor function. AMPK activation requires phosphorylation by the tumour suppressor protein liver kinase B1 (LKB1). In contrast, Akt can phosphorylate and inhibit the activity of AMPK. The LKB1 serine/threonine kinase is encoded by the tumour suppressor gene, STK11/LKB1, which harbours germ-line mutations in the inherited cancer predisposition, Peutz-Jeghers syndrome, and somatic mutations in sporadic cancers10. The PI3K/Akt/mTOR pathway regulates levels of VEGF (vascular endothelial growth factor), an angiogenic stimulator, in many tumour cell lines and overexpression of mTOR leads to increased VEGF production11.
In the present study, we investigated expression profile of the tumour suppressor genes PTEN and LKB1, and important signaling elements of the PI3K/Akt/mTOR pathway in leiomyomas and adjacent normal myometrium with an aim to observe the status of this signaling cascade in leiomyomas in women of reproductive age and to explore the possibility of targeting this pathway for future therapeutic implications.
Material & Methods
In this matched controlled, prospective, observational study, pairs of uterine leiomyoma (UL) and matched myometrium (MM) tissues were obtained from 33 premenopausal women (mean age 40±5.4 yr, range 28-49 yr) who underwent hysterectomy for leiomyomas in the proliferative phase of the menstrual cycle, between January 2012 and February 2014, at the Department of Obstetrics and Gynecology, King George's Medical University, Lucknow, India. Of the 53 women evaluated, only 33 were in proliferative phase. The study protocol was approved by the Institutional Ethical Committee for Human Research and informed written consent was obtained from all participants. Preoperative diagnosis of leiomyomas was based on transvaginal ultrasongraphy findings. The women had not received any hormonal treatment during the six months prior to surgery. Patients with autoimmune disease, pelvic inflammatory disease, adenomyosis, dysfunctional uterine bleeding, polycystic ovarian syndrome, endometriosis and those who took non-steroidal anti-inflammmatory drugs (NSAIDS), gonadotropin releasing hormone (GnRH) agonists or steroids during the previous three months were excluded from the study.
The size of the tumours varied from 0.5 to 12 cm. One tumour was dissected from each uterus. To maintain uniformity, leiomyomas selected for the study were 3.5 to 4.0 cm in diameter and were taken below the capsule. Myometrial samples, for use as paired controls were obtained from surrounding normal myometrium situated more than two cm away from the fibroid capsule. Each tissue sample was divided into two. One half was placed in 10 per cent neutral buffered formalin for histopathological examination and the other half in TRIzol reagent (Invitrogen, Carlsbad, CA, USA) for molecular analysis. The tissue for RNA extraction was stored at -80°C until use. After fixation for 24 h in formaldehyde the tissue was embedded in paraffin blocks at 56°C. All leiomyoma and MM samples were confirmed histologically (Fig. 1A, B).
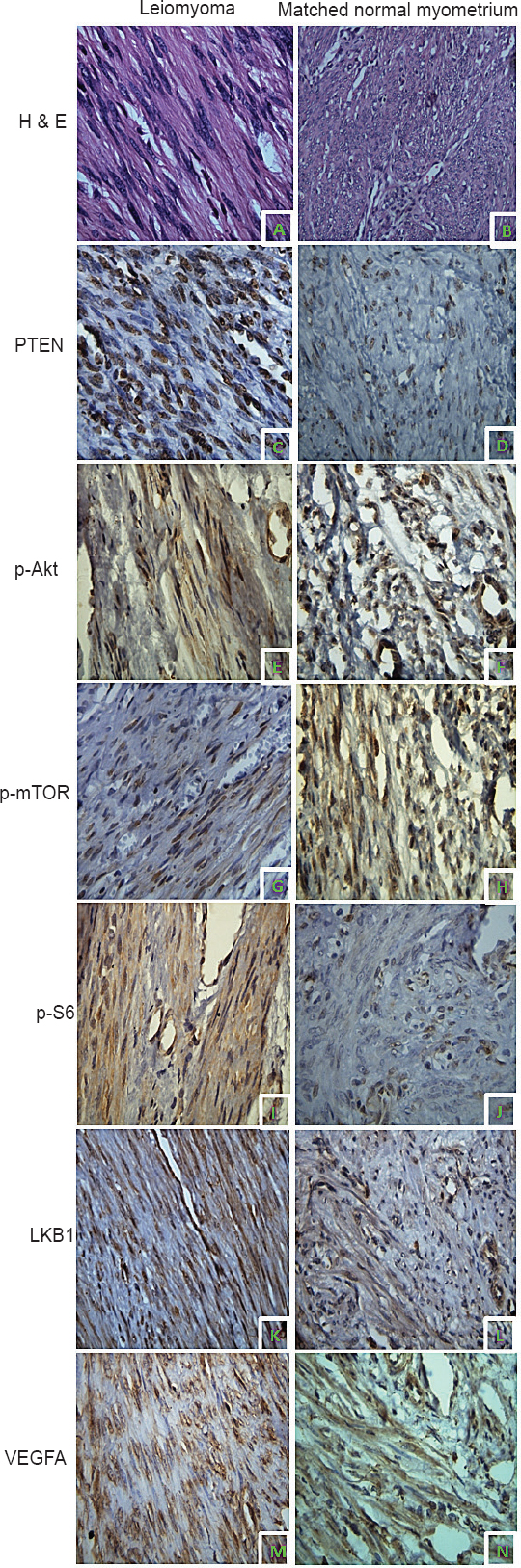
- Immunolocalization PI3K/Akt/mTOR pathway proteins in uterine leiomyoma (UL) and matched myometrium (MM). Hematoxylin and Eosin (H&E) stained section of UL (A) and MM (B). Phosphatase and tensin homolog deleted on chromosome ten (PTEN) expression in UL (C) and MM (D). pAkt expression in UL (E) and MM (F). Mammalian target of rapamycin phosphorylated at serine 2448 (p-mTOR) expression in UL (G) and MM (H). S6 ribosomal protein phosphorylated at serine 235/236 (pS6) expression in UL (I) and MM (J). liver kinase B1 (LKB1) expression in UL (K) and MM (L). Vascular endothelial growth factor A (VEGFA) expression in UL (M) and MM (N). Except for panel B (20x magnification), images in all other panels are captured with 40x magnification.
RNA isolation and quantitative real-time PCR (qPCR): Total RNA was extracted from UL and MM tissues with TRIzol reagent (Invitrogen, Carlsbad, CA, USA) following the standard manufacturer's protocol. RNA purification was done by DNase1 (Amplification grade; Invitrogen, Carlsbad, California, USA) treatment. RNA was quantified on a Picodrop spectrophotometer (Picodrop, UK) at 260/280 nm wavelength. cDNA synthesis was carried out starting with 250 ng total RNA using random hexamer primers with high capacity cDNA Reverse Transcription Kit (Applied Biosystems, CA, USA) as per manufacturers instruction. qPCR was performed in the presence of SYBR Green fluorescent dye using StepOne Real time PCR system (Applied Biosystems, CA, USA). Briefly, the reaction mixture consisted of reverse transcribed cDNA, 2× POWER SYBR Green master mix (Applied Biosystem, CA, USA), and 10 μM forward and reverse primers. β-actin, a housekeeping gene, was used as a normalization control. All PCR primers were synthesized by MWG Operon, India, and cross-checked by Primer Express software 3.0 (Applied Biosystems) and Blast sequence analysis (NCBI, USA). The sequences of all primers used in this study are listed in Table I 12131415. Thermal cycling conditions were 10 min at 95°C, followed by 40 cycles of 15 sec at 95°C and 1 min at 60°C. A negative control without template was run in parallel to assess the overall specificity of the reaction. Melt curve analysis was employed at the end of each PCR to confirm the specificity of the PCR product. Each measurement was performed in triplicate. Relative gene expression was determined by the ∆∆CT method, normalized to β-actin and expressed as fold change from MM set at a value of 1.0.

Immunohistochemistry: Paraffin-embedded tissue blocks of UL and MM were sectioned at 4 μm and mounted on APS (3-aminopropyl triethoxysilane) coated glass slides. Slides were incubated at 60°C for 1-2 h and then deparaffinized and rehydrated. Antigen retrieval was performed at 98°C for 15 min in a microwave oven (EZ antigen retriever system; BioGenex, USA) in Tris-EDTA buffer (pH 9.0) for antibodies against PTEN (Clone 138G6; Cell Signaling Technology, Boston, MA, USA), phospho-Akt (ser 473; Santa Cruz Biotechnology, Santa Cruz, CA, USA), phospho-mTOR (ser 2448; Santa Cruz Biotechnology), phospho-S6 (ser235/236; Cell Signaling Technology), LKB1 (Abcam, Cambridge, MA, USA) and VEGF-A (A-20; Santa Cruz Biotechnology). Sections were incubated with primary antibody in a humid chamber for 18 h at 4°C. Detection was performed using Novolink Min Polymer Detection System (Novacastra, Leica Biosystem Newcastle Ltd., UK). Visualization of immunoreactivity was achieved using 3, 3’-diaminobenzidine tetrahydrochloride as chromogen followed by hematoxylin counterstain. Slides were dehydrated with a series of graded ethanol washes and xylene washes, mounted with DPX (di-n-butylphthalate in xylene) mountant (Rankem, India) and cover-slipped for bright field microscopy. NIS-Elements (Nikon, Instruments Inc., Japan) software was used for image acquisition. Appropriate positive controls were selected according to the manufacturer's recommendations. Slides processed without the primary antibodies were used as negative controls.
All samples were anonymized and independently scored by two observers. In case of disagreement, the slides were re-examined and a consensus was reached by the observers. Scoring was semiquantitative using the Quickscore method16 for the intensity and proportion of cells stained for each marker. The immunointensity was scored on a scale of 0: no staining; 1: light staining; 2: moderate staining; 3: strong staining. The proportion of staining was scored on a scale of 1 : 0 to 4 per cent; 2 : 5 to 20 per cent; 3 : 21 to 40 per cent; 4 : 41 to 60 per cent; 5 : 61 to 80 per cent; and 6 : 81 to 100 per cent. Immunointensity and immunopositivity scores were multiplied giving results that ranged from 0 to 18. The final evaluation assigned samples as negative for sections with a score of 0-3, and positive for sections with a score ≥4.
Statistical analysis: All analyses were carried out using SPSS version 17.0 statistical software (SPSS Inc: Chicago, USA). The quantitative real time PCR results were evaluated by Student's t test. Immunohistochemical scoring was analyzed using sign test to determine appropriate cut-off for positive versus negative immunoexpression. The results are expressed in medians and percentages. The medians of positive scores (≥ 4) of various proteins in leiomyoma and their paired myometrium were compared by Wilcoxon signed-rank test. Odds ratios and 95% confidence intervals were also calculated.
Results
Gene expression in uterine leiomyoma and matched myometrium: Expression of PTEN, Akt1, Akt2, mTOR, LKB1 and VEGFA genes were assessed in uterine leiomyoma and matched myometrial tissues using real time PCR (Fig. 2). mRNA levels of PTEN (2.52 fold; P=0.03) and LKB1 (3.93 fold; P=0.01) were significantly higher in leiomyoma as compared to myometrium. VEGFA mRNA levels in leiomyoma were significantly downregulated compared to myometrium (2.95 fold; P=0.01). No significant change in the expression of Akt1, Akt2 and mTOR genes was found between UL and MM.
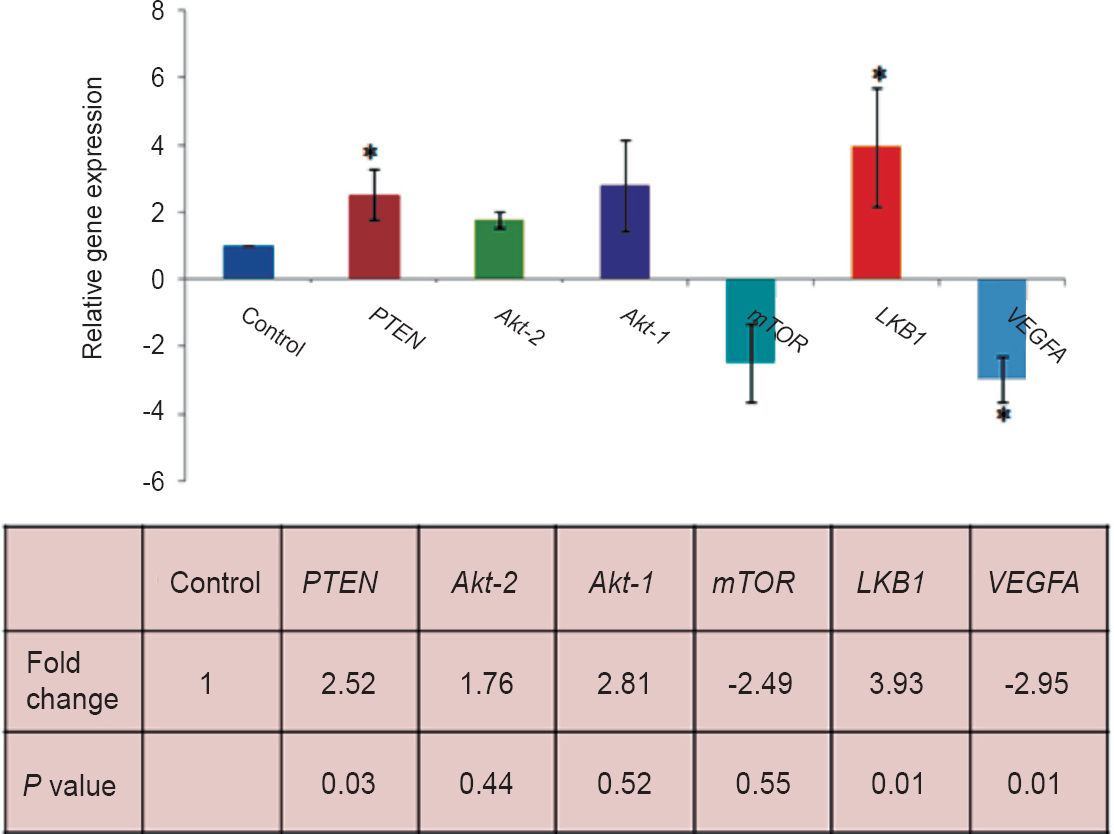
- Analysis of relative levels of PTEN, Akt1, Akt2, mTOR, LKB1 and VEGFA gene expression by real time PCR in uterine leiomyoma (UL) and matched myometrium (MM). Data were analyzed by the ΔΔCT method, normalized to β-actin and expressed as fold change from MM set at a value of 1.0. P values were calculated by Student t test.
Semiquantitative assessment of expression of various proteins in uterine leiomyomas and matched myometrium: Immunohistochemical staining was performed for six key proteins of the PI3K/Akt/mTOR signaling pathway: PTEN, phospho-Akt ser473 (pAkt), phospho-mTOR (p-mTOR), phospho-S6 (pS6), LKB1 and VEGFA. Chromogenic signals and cellular expression pattern were evaluated for each marker. Immunostaining for PTEN was primarily nuclear (Fig. 1C, D). Positive PTEN expression was observed in 94 per cent (31/33) of UL and 61 per cent (20/33) of MM (Table II). A comparison of immunoscores demonstrated a significantly higher PTEN expression in UL as compared to MM (P=0.015) confirming our real time findings.
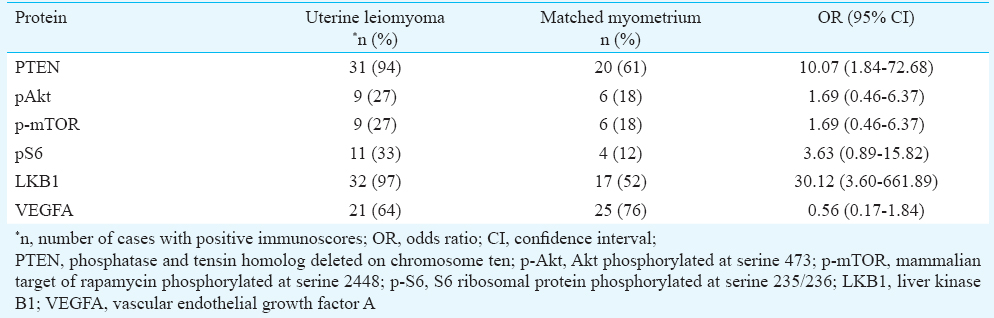
The activation status of Akt and mTOR was evaluated immunohistochemically by use of phosphospecific antibodies. Activity of the mTOR pathway was further assessed by detecting expression of phosphorylated S6 ribosomal protein. Immunohistochemical staining for pAkt (Fig. 1E, F), p-mTOR (Fig. 1G, H), and pS6 (Fig. 1I, J) was both cytoplasmic and nuclear. pAkt expression was seen in 27 per cent (9/33) leiomyoma and 18 per cent (6/33) myometrial tissues. Immunopositivity for p-mTOR and pS6 was observed in 27 per cent (9/33) and 33 per cent (11/33) of leiomyomas, respectively (Table II). In the MM group, 18 per cent (6/33) myometrium samples showed positive staining for p-mTOR and 12 per cent (4/33) were positive for pS6. The quickscore values in the leiomyomas compared to myometrial samples were not significantly different for all the three phosphoproteins (Table III). Immunostaining for the pAkt, p-mTOR, and pS6 in UL was localized mainly to the peripheral portions of the tumours.
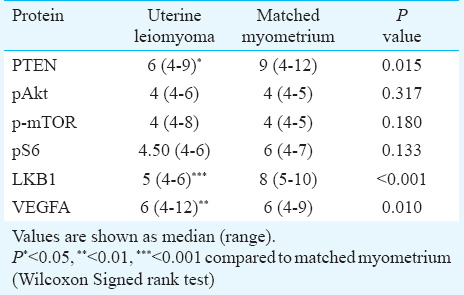
Immunohistochemical staining for LKB1 was predominantly cytoplasmic and less frequently nuclear (Figs. 1K, L). Positive LKB1 staining was observed in 97 per cent (32/33) of UL as compared to 52 per cent (17/33) MM (Table II) and the difference in immunoscores for LKB1 between the two tissues was significant (P<0.001) (Table III). A leiomyoma associated increase in LKB1 immunoreactivity confirmed our gene expression results.
The cellular immunolocalization of VEGFA was cytoplasmic (Figs. 1M, N). Positive VEGFA staining was observed in 76 per cent (25/33) MM and 64 per cent (21/33) UL, respectively (Table II). A paired comparison between the two tissues revealed a significantly lower VEGFA expression in UL as compared to MM (P=0.01) (Table III).
Discussion
In the present study, an upregulated expression of PTEN both at transcript and protein levels in UL was observed as compared to adjacent myometrial tissue. These findings corroborated with earlier studies1718 though either no change19 or lower20 PTEN expression in UL as compared to MM has also been reported. This could possibly be due to methodological differences and genetic variability of the study population.
Akt exists in mammalian cells as three highly conserved isoforms. While Akt1 and Akt2 are ubiquitously expressed, Akt3 displays a more restricted distribution and is found abundantly in nervous tissue21. We analysed mRNA levels of Akt1 and Akt2 as well as immunoexpression of pAkt (the activated form of Akt phosphorylated at Serine-473) in UL and MM. No significant difference was observed in the expression of these pro-survival factors between UL and MM. Similar pAkt expression in UL and normal myometrial tissue has been reported by other researchers222324. Further, pAkt expression was low or absent in a large number of leiomyoma samples (73%) in our study, a finding supported by another study25. Considering the fact that PTEN regulates Akt activation, low pAkt and increased PTEN levels observed in the present study suggest downregulation of the PI3K/Akt pathway in UL. Contrary to our findings, high levels of pAkt, either exclusively in leiomyomas from the growing period of tumours26 or in large fibroids27 have been reported. Aberrant expression of G-protein coupled receptor 10 (GPR10) has been shown to result in the activation of the PI3k/Akt pathway in uterine leiomyomas28. The disparity between the findings may be attributed to differences in sample selection criteria, hormonal milieu at the time of sampling, size of fibroids, age of patients, and/or ethnicity.
mTOR is a critical downstream effector of the PI3K/Akt pathway and phosphorylation of the ribosomal protein S6 is considered a good indicator of mTOR activation8. No significant difference was observed in mTOR mRNA and immunoscores for p-mTOR and pS6 between UL and MM. Low immunoscores for p-mTOR and pS6 protein as observed in majority of UL might be indicative of downregulation of mTOR pathway in these tumours. Considering that tuberin acts as a negative regulator of the mTOR pathway, we investigated its immunoexpression in UL and MM. Interestingly, the expression of tuberin was substantially higher in UL as compared to MM (data not shown), supporting the observation on downregulation of mTOR pathway in UL. These findings are also supported by other studies2325. Crabtree et al29 have shown upregulation of mTOR pathway in UL based on a global transcriptional analysis. Their results however, were inconclusive at protein level (S6 phosphorylation).
Immunostaining for pAkt, p-mTOR and pS6 in leiomyomas was localized mainly to the peripheral portions of the tumour. Pertinently, peripheral parts of leiomyomas have been shown to have a higher turnover of cells compared with the central parts30. In addition to pAkt, the tumour suppressor LKB1 also regulates the mTOR pathway. We observed increased mRNA and protein expression of LKB1 in UL as compared to MM. A previous study has shown activation of AMPK signaling pathway in primary ULs from Eker rats31. We found immunostaining for LKB1 predominantly in the cytoplasm of both UL and MM. The cytoplasmic portion of LKB1 has been reported to be responsible for cell cycle arrest and tumour suppressive activities32.
Loss of LKB1 confers an increased angiogenic potential in certain cell types by upregulation of VEGF33. We observed significant downregulation of VEGFA both at transcript and protein levels in UL as compared to MM. Conflicting results have been reported in literature on the expression of VEGFA3435. In one study, reduction in the content of VEGFA in large UL in comparison to small myomas and control myometrium was reported36. Authors have suggested that VEGFs, in contrast to their role in malignant tumours, do not participate in myometrium-myoma conversion and in myoma progression36. Consistent with upregulation of LKB1 expression observed in the present study, downregulation of VEGFA further supports the tumour suppressive role of LKB1 in UL.
In conclusion, our results demonstrated significantly higher levels of PTEN and LKB1 in UL as compared to MM. Increased expressions of these tumour suppressors in concert with negative or low levels of activated Akt, mTOR and S6 observed in majority of UL suggest downregulation of PI3K/Akt/mTOR pathway in these tumour contributing to their benign nature. Hence, this pathway alone may not play a significant role in pathogenesis of UL. Considering the fact that PI3K/Akt/mTOR pathway is influenced by feedback mechanisms and cross-talk with members of other well recognized signal transduction pathways (such as MAPK/ERK, Wnt/β-catenin and TGF-β signaling pathways), future studies involving simultaneous targeting of multiple pathways would be beneficial in the elucidation of molecular mechanisms underlying pathogenesis of leiomyoma with future therapeutic implications.
Acknowledgment
The authors acknowledge the technical assistance provided by Ms. Nidhi Verma and Shri. Kamlesh Sharma, and thank Drs Geetanjali Mishra and Abnish Gautam for their invaluable inputs in this study. Department of Biotechnology, New Delhi (Grant No. BT/PR5242/MED/30/803/2012) and Department of Science and Technology, New Delhi, India (Grant No.SR/WOS-A/LS-118/2008) are acknowledged for financial support.
Conflicts of Interest: None.
References
- Uterine fibroids: Clinical manifestations and contemporary management. Reprod Sci. 2014;21:1067-92.
- [Google Scholar]
- Complex networks of multiple factors in the pathogenesis of uterine leiomyoma. Fertil Steril. 2013;100:178-93.
- [Google Scholar]
- Sex steroidal regulation of uterine leiomyoma growth and apoptosis. Hum Reprod Update. 2004;10:207-20.
- [Google Scholar]
- Progesterone signaling in breast and endometrium. J Steroid Biochem Mol Biol. 2006;102:2-10.
- [Google Scholar]
- Effect of estrogen and inhibition of Phosphatidylinositol-3 kinase on Akt and FOXO1 in rat uterus. Steroids. 2007;72:422-8.
- [Google Scholar]
- Targeting the PI3K-Akt-mTOR signaling network in cancer. Chin J Cancer. 2013;32:253-65.
- [Google Scholar]
- Uterine-specific loss of Tsc2 leads to myometrial tumors in both the uterus and lungs. Mol Endocrinol. 2013;27:1403-14.
- [Google Scholar]
- PI3K-Akt and MAP kinase signaling pathways in polycystic ovarian syndrome, uterine leiomyomas and endometriosis: an update. Gynecol Endocrinol. 2012;28:175-81.
- [Google Scholar]
- Targeting the PI3K-Akt pathway in human cancer: rationale and premise. Cancer Cell. 2003;4:257-62.
- [Google Scholar]
- The LKB1-AMPK pathway: metabolism and growth control in tumor suppression. Nat Rev Cancer. 2009;9:563-75.
- [Google Scholar]
- Targeting mTOR pathway: a new concept in cancer therapy. Indian J Med Paediatr Oncol. 2010;31:132-6.
- [Google Scholar]
- Rapamycin and CCI-779 inhibit the mammalian target of rapamycin signaling in hepatocellular carcinoma. Liver Int. 2010;30:65-75.
- [Google Scholar]
- Somatic mutations of the Peutz-Jeghers syndrome gene, LKB1/STK11, in malignant melanoma. Oncogene. 1999;18:1777-80.
- [Google Scholar]
- VEGF-A immunohistochemical and mRNA expression in tissues and its serum levels in potentially malignant oral lesions and oral squamous cell carcinomas. Oral Oncol. 2012;48:233-9.
- [Google Scholar]
- Molecular evidence for two forms of Crohn disease. Proc Natl Acad Sci USA. 1994;91:12721-4.
- [Google Scholar]
- A “quickscore” method for immunohistochemical semiquantitation: validation for oestrogen receptor in breast carcinomas. J Clin Pathol. 1995;48:876-8.
- [Google Scholar]
- Sphingolipid content in the human uterus and pair-matched uterine leiomyomas remains constant. Lipids. 2013;48:245-50.
- [Google Scholar]
- 17β-estradiol induces up-regulation of PTEN and PPARγ in leiomyoma cells, but not in normal cells. Int J Oncol. 2010;36:921-7.
- [Google Scholar]
- Phosphorylation of PTEN (phosphatase and tensin homologue deleted on chromosome ten) protein is enhanced in human fibromyomatous uteri. J Steroid Biochem Mol Biol. 2007;103:196-9.
- [Google Scholar]
- Expression of proliferative and preapoptotic molecules in human myometrium and leiomyoma throughout the menstrual cycle. Reprod Sci. 2007;14:678-86.
- [Google Scholar]
- The emerging role of the phosphatidylinositol 3-kinase/Akt/mammalian target of rapamycin signaling network in cancer stem cell biology. Cancer. 2010;2:1576-96.
- [Google Scholar]
- Downregulation of the integrin αv signaling pathway in uterine leiomyomas. Gynecol Obstet Invest. 2011;71:129-35.
- [Google Scholar]
- Increased expression of tuberin in human uterine leiomyoma. Fertil Steril. 2011;95:1805-8.
- [Google Scholar]
- Differential expression of receptor tyrosine kinases (RTKs) and IGF-1 pathway activation in human uterine leiomyomas. Mol Med. 2008;14:264-75.
- [Google Scholar]
- Constitutive activation with overexpression of the mTORC2-phospholipase D1 pathway in uterine leiomyosarcoma and STUMP: morphoproteomic analysis with therapeutic implications. Int J Clin Exp Pathol. 2011;4:134-46.
- [Google Scholar]
- Differential expression of Akt/protein kinase B, Bcl-2 and Bax proteins in human leiomyoma and myometrium. J Steroid Biochem Mol Biol. 2003;87:233-40.
- [Google Scholar]
- Expression of insulin-like growth factors (IGFs) and IGF signaling: molecular complexity in uterine leiomyomas. Fertil Steril. 2009;91:2664-75.
- [Google Scholar]
- Loss of repressor REST in uterine fibroids promotes aberrant G protein-coupled receptor 10 expression and activates mammalian target of rapamycin pathway. Proc Natl Acad Sci USA. 2013;110:2187-92.
- [Google Scholar]
- Comparison of human and rat uterine leiomyomata: identification of a dysregulated mammalian target of rapamycin pathway. Cancer Res. 2009;69:6171-8.
- [Google Scholar]
- Different proliferative and apoptotic activity in peripheral versus central parts of human uterine leiomyomas. Gynecol Obstet Invest. 2003;55:199-204.
- [Google Scholar]
- Stromal liver kinase B1 [STK11] signaling loss induces oviductal adenomas and endometrial cancer by activating mammalian target of rapamycin complex 1. PLoS Genet. 2012;8:e1002906.
- [Google Scholar]
- Akt blocks the tumor suppressor activity of LKB1 by promoting phosphorylation-dependent nuclear retention through 14-3-3 proteins. Am J Transl Res. 2012;4:175-86.
- [Google Scholar]
- Vascular abnormalities and deregulation of VEGF in LKB1-deficient mice. Science. 2001;293:1323-6.
- [Google Scholar]
- Quantification of vascular endothelial growth factor-A in leiomyomas and adjacent myometrium. Clin Sci. 2001;101:691-5.
- [Google Scholar]
- [Leiomyoma ve normal myometrium da immunohistokimyasalvaskulerendotelyal growth factor (VEGF) ekspresyonu] Turk Fertilite Dergisi. 2003;11:299-304.
- [Google Scholar]
- Does vascular endothelial growth factor participate in uterine myoma growth stimulation? Eur J Obstet Gynecol Reprod Biol. 2012;164:93-7.
- [Google Scholar]




