Translate this page into:
Sialic acids siglec interaction: A unique strategy to circumvent innate immune response by pathogens
Reprint requests: Dr Chitra Mandal, Cancer Biology & Inflammatory Disorders Division, CSIR-Indian Institute of Chemical Biology, 4, Raja S.C. Mullick Road, Kolkata 700 032, India e-mail: chitra_mandal@yahoo.com or cmandal@iicb.res.in
-
Received: ,
This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-Share Alike 3.0 Unported, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This article was originally published by Medknow Publications & Media Pvt Ltd and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Sialic acids (Sias) are nine-carbon keto sugars primarily present on the terminal residue of cell surface glycans. Sialic acid binding immunoglobulins (Ig)-like lectins (siglecs) are generally expressed on various immune cells. They selectively recognize different linkage-specific sialic acids and undertake a variety of cellular functions. Many pathogens either synthesize or acquire sialic acids from the host. Sialylated pathogens generally use siglecs to manipulate the host immune response. The present review mainly deals with the newly developed information regarding mechanism of acquisition of sialic acids by pathogens and their biological relevance especially in the establishment of successful infection by impairing host innate immunity. The pathogens which are unable to synthesize sialic acids might adsorb these from the host as a way to engage the inhibitory siglecs. They promote association with the immune cells through sialic acids-siglec dependent manner. Such an association plays an important role to subvert host's immunity. Detailed investigation of these pathways has been discussed in this review. Particular attention has been focused on Pseudomonas aeruginosa (PA) and Leishmania donovani.
Keywords
Elastase
innate immunity
Leishmaniasis
neutrophil extracellular traps (NETs)
Pseudomonas aeruginosa
reactive oxygen species
Sialic acids
Siglecs

Introduction
Structural diversity and unique strategic location of sialic acids (Sias) on the cells make them one of the most important molecules in life and set the challenges for sialoglycobiologist1. Sialic acids are nine carbon acidic sugars typically found as the terminal residue of cell surface sugar chains as well as on secreted glycoproteins and in the extracellular matrix. About 50 different modifications exist for sialic acids in nature2.
Sialic acids possess contrasting character of working as masking element of an array of cell surface receptors whereas on the other side these function as a recognition site for various lectins and antibodies, indicating the unique nature of the molecule2345. Many immunological functions are attributed to sialic acids such as formation of negatively charged barrier for host to reduce interaction with pathogen and dampening the classical or alternative pathways of complement by selective deposition on pathogen surface, thus functioning as both beneficial as well as harmful for the system6789.
Sialic acid binding immunoglobulins (Ig)-like lectins (siglecs) belong to I-type lectin with a selective expression on the haematopoetic cell lineages. These have amazing structural diversity to recognize and interact with an array of linkage-specific sialic acids on a glycan structure express on host cells as well as pathogen10. Fourteen human and nine murine siglecs have already been identified and the list is still increasing11.
Based on the inter-species sequence similarity and conservation in mammals, siglecs are mainly classified into two families. Siglec-1 (sialoadhesin or CD169), siglec-2 (CD22), siglec-4 (myelin-associated glycoprotein, MAG) and siglec-15 are evolutionary conserved with poor inter-species sequence similarity. Most of the mammalian species contain definite orthologue of all these siglecs. Other family of siglecs, mainly siglec-3/CD33 related siglecs has encountered rapid evolution indicated by variable interspecies sequence homology making it difficult to find orthologue among species1213. These show 50-99 per cent sequence homology. It comprises the largest family containing 10 human (siglec-3, -5, -6, -7, -8, -9, -10, -11, -14, -16) and five rodent siglecs (siglec-3, -E, -F, -G, -H)12.
Siglec-3/CD33 related-siglecs express mostly on haematopoetic cell lineages. Siglec-9 is expressed on neutrophils, monocytes, fraction of natural killer (NK) cells, B cells etc., while siglec-8 appearance is restricted on the circulating eosinophils and negligible on besophils141516. Several siglecs can be present on the same cells, such as monocytes express siglec-3, -5, -7, -9 and -10 indicating the extent of functional redundancy at cellular level17181920. However, siglec-4 is expressed only on neuronal cells.
After binding with terminally sialylated glycoconjugates, siglecs undertake various functions such as internalization of sialylated pathogens, attenuation of inflammation, restraining cellular activation, attenuation of damage-associated molecular pattern-mediated inflammation along with inhibition of NK cell activation2122.
Innate immune system is the first line of defense evolved during the years of evolution. This is responsible for controlling and clearing invading pathogens by non specific means such as toll like receptors (TLRs), complement or by selectively activating the adaptive immune response232425. Polymorphonuclear neutrophils (PMN) are crucial innate immune cells that protect host from invading bacterial and fungal infections by using different defense strategies. Neutrophils exhibit phagocytic activity against pathogenic bacteria and release antimicrobial molecules. Several potent antimicrobial molecules like cationic peptides, proteases, lactoferrin, reactive oxygen species (ROS) and reactive nitrogen intermediates are produced by the neutrophils to serve as potent innate immune cells against pathogens26. Neutrophils can also combat severe infection by neutrophil extracellular traps (NETs) formation, a mechanism by which activated neutrophils eliminate pathogenic bacteria. Macrophages, another innate immune cells, are the professional antigen presenting cells (APCs) found nearly in all tissues, to effectively present antigen to the T cells27.
Although siglecs were identified in early eighties, not much work has been done to explore the consequences of sialic acids acquisition by various pathogens and their interactions with immune system in sialic acids-siglec dependent manner in promoting infection or mediating immune activation.
This review highlights the sialic acids-siglec interaction-mediated innate immune escape of pathogens with special reference to an opportunistic Gram-negative bacteria Pseudomonas aeruginosa (PA). Our group has demonstrated the status of sialic acids on PA and their role in host recognition through involvement of human siglecs present on the immune cells28. Sialic acids-mediated interactions of PA with siglec-9 present on neutrophils dampen the innate immune functions29. Such an interaction during Leishmania donovani infection has also been discussed briefly.
Detection of sialic acids on pathogens
Considering the vast microbial kingdom, assessment of the sialoglycan profile remains a relatively unexplored domain of microbial sialobiology. Although a few sialylated microorganisms have been reported (Table I)2829303132333435363738394041424344454647484950515253545556, sialylation status on PA remains untouched. Our group has recently demonstrated the status of sialic acids on PA by several analytical, biochemical and immunological methods2857. Sialic acid (Neu5Ac), its glycolyl derivative (Neu5Gc) and O-acetylated form (Neu5,9Ac2) on the surface of PA are detected by thin layer chromatography and fluorimetric-HPLC (Fig. 1A). The chromatogram exhibited well-resolved intense peaks corresponding to Neu5Ac, Neu5Gc and Neu5,9Ac2. Additionally, matrix-assisted laser desorption/ionisation-time of flight mass spectrometry (MALDI-TOF-MS) of purified Neu5Ac, Neu5Gc and Neu5,9Ac2 matched the expected signal having m/z at 448.7, 464.8 and 490.6, respectively confirming their occurrence on PA (Fig. 1B). Binding with Sambucus nigra agglutinin (SNA) and Maackia amurensis agglutinin (MAA) having specificity towards α2,6- and α2,3-linked sialic acids, respectively confirm the presence of linkage-specific sialic acids on PA5859 (Fig. 2). The bacterial membrane fraction showed stronger binding with SNA compared to Galanthus nivalis agglutinin, specific for terminal mannose (1,3), (1,6) and (1,2) mannose, Datura stramonium agglutinin, specific for galactose (1,4) N-acetyl glucosamine and peanut agglutinin, specific for galactose (1,3) N-acetyl galactosamine60. Further evidence for the presence of sialic acids is obtained using different siglecs where siglec-9 shows maximal binding with this bacteria33.
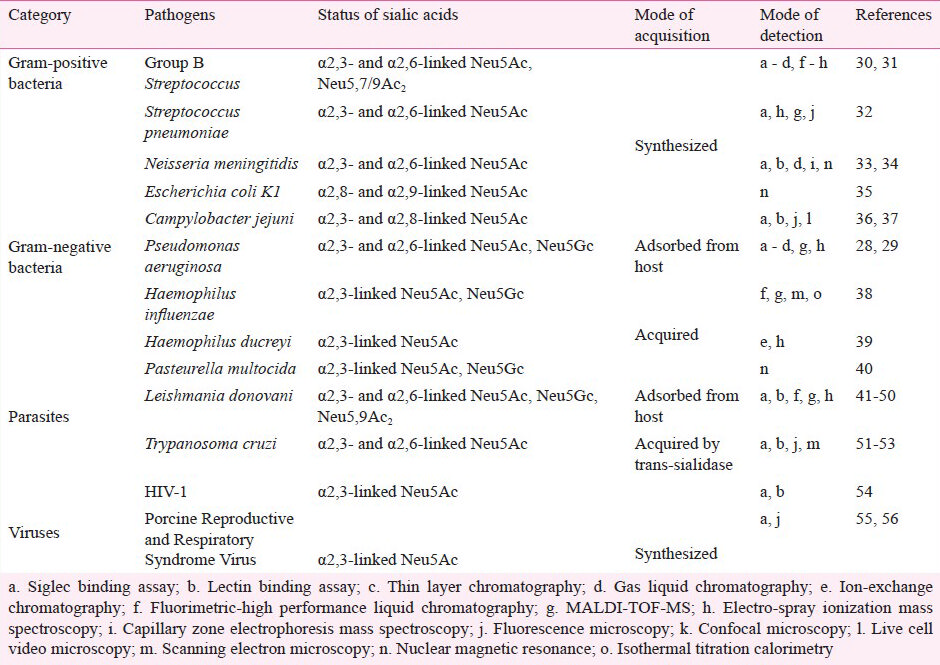
- Identification of sialic acids of Pseudomonas aeruginosa (PA) by analytical methods. (A) Fluorimetric-HPLC of 1,2-diamino-4,5-methyleneoxybenzene (DMB) derivatized sialic acids. A representative fluorimetric reverse phase HPLC profile of DMB derivatized sialic acids purified from PA. Three intense peaks were observed corresponding to Neu5Ac, Neu5Gc and Neu5,9Ac2. Reproduced with permission from John Wiley and Sons, Oxford, UK (FEBS Lett2010; 584: 555-61). B. MALDI-TOF-MS spectra of the total sialic acids from PA. Sialic acids were purified from PA+Sias and derivatized with 1,2-diamino-4,5-methyleneoxybenzene (DMB). These were analyzed by MALDI-TOF MS on a target with an equal volume of matrix (2,5-dihydroxybenzoic acid). All mass spectra were recorded in the positive ion mode. Sodium cationized molecular ions of Neu5Ac, Neu5Gc and Neu5,9Ac2 peaks having m/z at 448.7, 464.8 and 490.6, respectively are shown. Reproduced with permission from John Wiley and Sons, Oxford, UK (FEBS Lett 2010; 584: 555-61).
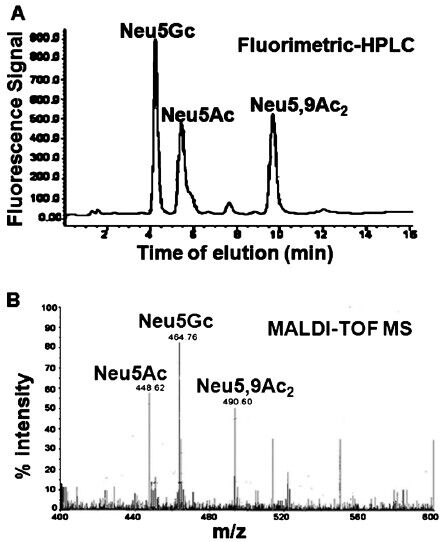
- Interaction of pathogens through linkage-specific sialic acids with different siglecs present on neutrophil and macrophages. The two pathogens namely L.donovani and P. aeruginosa contain α2,3 and α2,6 linked sialic acids on their surface. These linkage-specific sialic acids bind with siglec-1 and siglec-5 present on macrophages whereas siglec-5 and siglec-9 present on neutrophils.
PA also possesses α2,6-linked Neu5,9Ac2 on its surface as confirmed through binding with a lectin, Achatinin-H having preferential affinity towards Neu5, 9Ac2α2, 6GalNAc sialoglycotope42436162. A comparable amount of 9-O-acetylated sialic acids as a percentage of total sialic acids was detected on intact bacteria and its membrane fraction by fluorimetric estimation6364. To demonstrate this binding specificity toward the 9-O-acetyl moiety, bacteria were incubated with a recombinant acetylesterase to remove the O-acetyl group6566. The resultant de-O-acetylation caused a near total abolition of Achatinin-H binding, confirming the presence of linkage specific 9-O-acetylated sialic acids on PA.
By using similar analytical methods and probes, our group has also reported that both L. donovani promastigotes and amastigotes contain α2,6- and α2,3-linked sialic acids (Neu5Ac, Neu5Gc, Neu5,9Ac2) on their cell surface434445464748 (Fig. 2). Trypanosoma, another major genus of kinetoplastida, possesses highly sialylated and glycosylphosphatidylinositol (GPI) anchored dense mucin layer on its surface67.
Acquisition of sialic acids by pathogens
Detection of these sialic acids raises the obvious question regarding the mechanism(s) adopted by the pathogens to acquire these terminal sugar molecules. More than 20 pathogenic organisms either synthesize or acquire sialic acids from the host (Table I)282930313233343536373839404142434445464748495051525354555668. A few Gram-negative bacteria like Haemophilus influenza, Pasteurella multocida and H. ducreyi use their growth medium as a source of sialic acids whereas GBS, a Gram-positive bacteria and some others Gram-negative bacteria such as Escherichia coli K1, Neisseria meningitidis and Campylobacter jejuni have capability to synthesize sialic acids3031323334353637383940414243. The mechanism by which these sialic acids present in human serum are absorbed by PA remains to be explored.
Bioinformatic searches through the genome of PA indicate the absence of a defined endogenous biosynthetic pathway that prompted us to examine whether the sialic acids on the PA surface are derived from the growth medium. The medium (TSB) with 10 per cent heat inactivated normal human serum used for culturing PA (PA+Sias) showed the presence of Neu5Ac and Neu5Gc whereas the spent media demonstrated reduced levels of these sialic acids, suggesting possible adsorption of sialic acids from environment29. The decrease in the amount of sialic acids on PA+Sias with decreasing concentration of human serum in growth medium further suggested adsorption of sialic acids from culture medium (Fig. 3)29. We have not found any key enzyme in the sialic acids biosynthesis pathway in Leishmania. Moreover, trans-sialidase is also absent in this parasite. We have demonstrated that the sialic acids are adsorbed by promastigotes and amastigotes from the host4950.
![Adsorption of sialic acids by PA from normal human serum, used as a source of sialic acids. PA was cultured in tryptic soy broth (TSB) containing increasing concentrations of heat inactivated normal human serum (HI-NHS, 2-10%). Flow cytometric analysis showed adsorption of α2,6-linked sialic acids [Sambucus nigra agglutinin (SNA) positivity, filled square] and α2,3-linked sialic acids [Maackia amurensis agglutinin (MAA) positivity, filled circle] from HI-NHS. Data were presented as per cent of FITC-lectins positive PA cells. Reproduced with permission from Society for Leukocyte Biology, Bethesda, USA (J Leukoc Biol 2012; 91: 641-55).](/content/175/2013/138/5/img/IJMR-138-648-g004.png)
- Adsorption of sialic acids by PA from normal human serum, used as a source of sialic acids. PA was cultured in tryptic soy broth (TSB) containing increasing concentrations of heat inactivated normal human serum (HI-NHS, 2-10%). Flow cytometric analysis showed adsorption of α2,6-linked sialic acids [Sambucus nigra agglutinin (SNA) positivity, filled square] and α2,3-linked sialic acids [Maackia amurensis agglutinin (MAA) positivity, filled circle] from HI-NHS. Data were presented as per cent of FITC-lectins positive PA cells. Reproduced with permission from Society for Leukocyte Biology, Bethesda, USA (J Leukoc Biol 2012; 91: 641-55).
In this scenario, it may be envisaged that these sialic acids may initially be fragmented or degraded fully or partially by cellular enzymes and subsequently may be available for adsorption by pathogens. Adsorption of sialic acids from culture medium may be either by transglycosylation or by integration of sialic acids to the polyanionic lipophosphoglycan (LPG), LPG/proteophosphoglycan rich cell surface of pathogens. Another option is catalytic transfer of sialic acids from the nucleotide sugar donor CMP-Neu5Ac onto acceptor glycoconjugates on pathogens by serum sialyltransferase.
The presence of a unique enzyme trans-sialidase (TS) in trypanosome facilitates the transfer of glycosidically bound sialic acids from serum sialoglycoconjugates. Although Trypanosoma is devoid of any indigenous sialic acids biosynthesis machinery, trans-sialidase accomplishes the function of sialic acids acquisition. Trans-sialidase cleaves the sialic acids containing glycoconjugates from the host cell and transfers it to the parasite surface69. The uniqueness of this enzyme lies in that unlike sialyltransferase instead of transferring activated CMP-sialic acid, trans-sialidase transfers carbohydrate-linked sialic acids to the glycan structure and forms a new α2,3-glycosidic linkage to galactose or N-acetylgalactosamine707172.
Biological relevance of sialic acids on pathogens
The major immunological advantage of sialic acids acquisition by pathogens is to subvert the host immunity by acting as a molecular mimic as this sugar is the indispensable factor for the host73. One of the major functions of trans-sialidase is to subvert the complement deposition by acquiring sialic acids on parasite surface70. C3 is the central component of the complement system. C3b being a product of C3 activation, binds covalently through a reactive intramolecular thioester in the C3d domain to bacterial surfaces74. Due to the presence of sialylated lipooligosaccharide on the surface, both N. meningitidis and N. gonorrhoeae show limited C3-deposition on their surface75. In parallel, high sialic acids content of type III GBS strains also inhibits C5a production76. Interestingly, PA+Sias also shows lower anti-C3 binding compared to PA-Sias indicating a direct relationship between the sialic acids levels and C3-deposition on PA, suggesting a way of escaping from the host serum28. In contrast, enhanced expression of 9-O-acetylated sialoglycoproteins on erythrocytes of patients with visceral leishmaniais777879808182, increases the activation of alternative complement pathway-mediated haemolysis which leads to anaemia41. Along with this, sialic acids acquired by the parasite act as a virulent factor to help in dampening the immune response during infection4950.
Immunological relevance of siglecs
Innate immunity is body's first line of defense containing both humoral and cellular immune component. The major disadvantage in this process is the collateral damage of the host cells in due course. To minimize the damage of self, nature has evolved some host-specific ligands that engage some inhibitory receptors present on immune effector cells such as NK cells, neutrophils, monocytes and macrophages to restrict the magnitude of immune reaction. Siglecs mainly act as a negative regulator in immune cells by constraining the magnitude of immune reaction122425.
Siglec-mediated regulation of cellular activation
Most of the CD33-related siglecs posses a membrane proximal immunoreceptor tyrosine based inhibitory motif (ITIM) and a membrane-distal ITIM like motif12. ITIM motif is more dominant than ITIM like motif for recruitment of phosphatase and the phosphatase-mediated inhibitory functions83. When a sialylated ligand binds with the siglec, ITIMs carry out the signal to the downstream molecule. After phophorylation by Src-family kineses, ITIMs act as a docking site for binding of either Src homology 2 domain-containing inositol polyphosphate 5’ phosphatase (SHIP) or Src homology 2 domain-containing protein tyrosine phosphatases SHP-1 and SHP-2. These phosphates get activated and dephosphorylate an array of phosphorylated molecules, thus play a major role in balancing cellular response or controlling over-activation by antagonizing with the signals coming from immunereceptor tyrosine-based activation motifs (ITAMs)84. This pathway mainly has been explored in B- and NK cells84. Human siglec-14,-15, murine CD33 and siglec-H lack ITIM motif interact with DNAX activating protein of 12 kD (DAP12) which subsequently carry out the downstream signaling85. Siglec-H does not bind to sialic acids, thus cannot be designated as orthodox siglec86.
Cis-trans interaction
Sialic acids-siglec interaction is one of the crucial factors in balancing the positive and negative signals in a cell. Linkage specificity of sialic acids with the penultimate sugars, nature of side chain derivatives, structure of distant sugar residues and their modifications, etc. regulate the nature of the sialic acids-siglec interaction24. Siglecs on a cell make bonding with its own cell surface sialic acids named as cis interaction whereas during trans interaction, siglecs undergo molecular interaction with the sialic acids present on other cells12. Due to the high local concentration (˜100 mM in B cells) of sialic acids on the immune cell surface, most of the siglecs except sialoadhesin are masked by the sialic acids of its own cell in cis interaction8788. Protruded structure of sialoadhesin help them to stay much way out of cell surface and therefore making it unable to form cis interaction with the membrane sialic acids8990. During physiological condition, cis conformation of siglecs in a cell undergoes renovation into trans conformation in consequence of cellular activation or in vicinity with the sialylated pathogens. Presence of membrane sialidase or change in glycosylation pattern or reorganization of plasma membrane domain also regulate the conversion of cis interaction into trans91.
In resting B cell, inhibitory receptor siglec 2 (CD22) remains mostly in masked condition, and undergoes unmasking during its activation929394. Cis interaction has significant impact on modulating the cellular functions by inhibiting the non specific interaction of siglecs with any unintruded cell or sialylated molecules that can initiate unwanted signaling. However, cis and trans interaction in a cell is a dynamic process, trans interaction cannot totally preclude the cis interaction of a cell9596. NK cell inhibitory receptor namely siglec-7 having specificity for the Neu5Acα2,8Neu5Ac bearing glycotope shows little cytotoxicity on target cell due to masking effect caused by cis interaction. However, pre-treatment of NK cells with sialidase increase its cytotoxic activity due to the unmasking of siglec-7 from cis interaction9798.
Sialic acids-siglec interaction subverts innate immunity
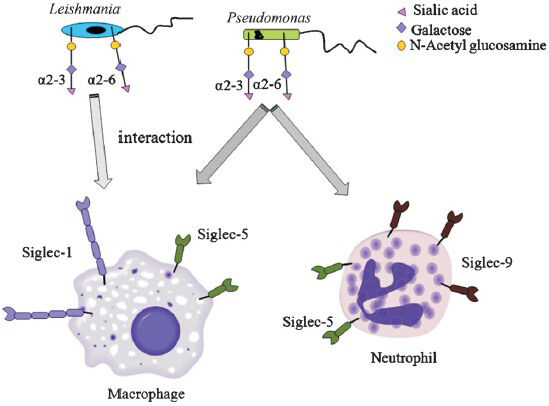
The persistence and successful establishment of pathogenic infection requires close interactions with host cells. The molecular determinants require for such interactions often include glycoproteins or glycolipids of the membrane. Siglecs on immune cells act as specific ligands for recognition mediated via sialic acids on several pathogenic bacteria333637. Porcine reproductive and respiratory syndrome virus uses sialic acids present on envelope for infecting pig alveolar macrophages through siglec-15599. N. meningitidis, C. jejuni, group B Streptococcus and T. cruzi also exhibit sialic acids-mediated interactions with siglec-1 and several CD33-related siglecs on host cell Table II28293032333652535456100101102.
Sialylated capsular polysaccharide (CPS) of various GBS serotypes specifically recognizes siglec-5,-7,-9 present on human leukocytes for establishment of successful infections30. Engagement of sialylated glycan of GBS serotype III through the siglec-9 of neutrophils subverts the host innate immune response by transmitting negative regulatory signals100. GBS Ia strains can also bind to human siglec-5 in a sialic acids-independent manner through a specific protein anchored to the bacterial cell wall and impairs immune evasion101. The specific engagement of sialylated lypo-oligosaccharides on C. jejuni with siglec-1 plays a crucial role in the pathogenesis of Guillain-Barre΄ syndrome102. Similarly, C. jejuni also interacts with siglec-7 and modulates inflammatory as well as immune responses of host (Table II)28293032333652535456100101102.
We have investigated the role of adsorbed sialic acids on PA in microbial pathogenesis through association with host cells. Binding of PA+Sias with recombinant soluble siglec-7/9 as well as CHO-siglec-7 and CHO-siglec-9 is considerably higher indicating the association of bacteria with innate immune cells through siglecs2829.
Co-infection is one of the major problems in most of the immune suppressive diseases. Co-infection with PA in some of the visceral leishmaniasis patients led us to carry forward the study of sialic acids-siglec interaction in the immune cells of these patients. Siglec-7 and siglec-9 expressing on CD56+ (NK cells) and CD14+ (monocytes) cells isolated from peripheral blood mononuclear cells of active visceral leishmaniasis patient showed higher binding with PA+Sias than those cells isolated from normal PBMC28. Thus, it may be envisaged that sialic acids-siglec interaction plays a significant role in establishing bacterial infection in immune compromised hosts.
Neutrophils express more siglec-9 and to some extent siglec-5 on its surface (Fig. 2). PA+Sias shows enhanced binding with neutrophils and sialidase-treated neutrophil compared to PA-Sias. Due to blocking of siglec-9 with anti-siglec-9 antibody on neutrophils, the binding of PA+Sias is reduced indicating direct involvement of siglec-9. In contrast, antibody against siglec-5-treated neutrophils shows no such decrease in binding recommending adhesion of PA with neutrophils maximally through sialic acids-siglec-9 interaction29.
Attenuation of oxidative burst of neutrophils by PA+Sias
During ingestion of microorganisms, neutrophils undergo “respiratory burst” leading to striking increase in oxygen consumption103. Neutrophils possess NADPH oxidase which is a complex of membrane-bound multi-component enzyme. Different bacteria/immune complexes after association with host cell-surface receptors release soluble mediators (chemoattractant peptides and chemokines) which can activate NADPH oxidase quickly. The activated enzyme releases electron from NADPH. The major intermediate for such reaction is superoxide anions O2−·. Dismutation of superoxide subsequently produces hydrogen peroxide (H2O2). H2O2 converts nontoxic compounds into highly reactive, toxic metabolites and then these become antimicrobial. Additionally, hydroxyl radical (·OH) and hypochlorous acids (HOCl) are also formed88. Myeloperoxidase, a granular enzyme, catalyses to produce HOCl from Cl−. This enzyme is mainly expressed in immature and mature neutrophils and also in monocytes104. Thus activated NADPH oxidase creates large quantities of ROS in the phagosome of neutrophil which combat with invading pathogens.
Due to interaction with PA+Sias through sialic acids-siglec-9 neutrophil produces comparable amount of ROS as that of untreated neutrophil suggesting functional impairment of host's first line of defense. In contrast, due to absence of sialic acids in PA-Sias, the interaction of sialic acids-siglec-9 is inhibited. As a result, higher amount of ROS is generated by the neutrophils thereby inhibiting bacteria to establish infection in host29.
Due to reduced or absence of oxidative burst of neutrophil, chronic granulomatous disease patients are frequently infected with an array of microbial pathogens including Staphylococcus, Salmonella, Aspergillus, and Candida species to spread out lymphadenitis, skin abscesses and pneumonia105106.
Interaction of siglec-9 with sialic acids on PA+Sias facilitates reduced release of elastase from neutrophil
Leukocyte granule–associated proteinase such as neutrophil elastase is a 30-kD glycoprotein with potent catalytic activity dictated by a catalytic triad that consists of histidine, asparagine and serine residues by forming a charge-relay system107. Elastase presents at high concentrations in the neutrophil granules. It acts as an antimicrobial agent by disrupting the structural integrity of invading microbes. Toxins of Shigella, Salmonella, and Yersinia are efficiently degraded by neutrophil elastase. This enzyme effectively kills them in its phagosomes108.
Due to the engagement of PA+Sias to neutrophil through sialic acids-siglec-9 interaction, the release of elastase is significantly reduced with respect to PA-Sias. Blocking of sialic acids-siglec-9 interaction either by specific antibody or removal of sialic acids, innate immune cells secrete significantly higher amount of elastase signifying importance of this interaction in impairment of the neutrophil's function29. Outer membrane protein A of E. coli, K. pneumonia and PA strain (H103) degrade neutrophil elastase to establish their infection. Moreover, ecotin, a serine protease inhibitor of E. coli shows a protective activity against neutrophil elastase109.
Engagement of sialic acids and siglec-9 attenuates the neutrophils extracellular traps formation by neutrophil
Neutrophils extracellular traps (NETs) are composed of extracellularly released chromatin material, serine proteases and cytoplasmic proteins27. Elastase, cathepsin G, azurocidin and proteinase 3 are the main serine proteases present in neutrophils and play essential role in acute infections and inflammation. Due to these high serine proteases, both Gram-negative as well as Gram-positive bacteria and their virulent proteins are degraded extracellularly by NETs110.
Production of NETs as shown by clear network of extracellular fibers is attenuated after engagement of neutrophils with PA+Sias through siglec-9-sialic acids association (Fig. 4). Thus PA+Sias shows resistance towards NETs-mediated killing by neutrophils demonstrating impairment of the power of innate immune cells for the effective clearance of bacteria due to Sias-siglec-9 interaction. In contrast, enhancement of such extracellular fibers is observed after engagement of neutrophil with PA-Sias or sialidase-treated PA+Sias due to minimal siglec-9-sialic acids interaction to eliminate the bacteria29.
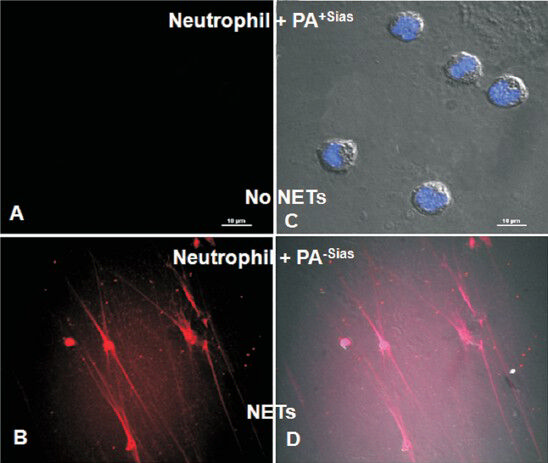
- Visualization of neutrophil extracellular traps (NETs) by confocal microscopy. PA was incubated with neutrophils. Neutrophil extracellular traps (NETs) were visualized under confocal microscopy after stained with Sytox orange (DNA stain for dead cell) only as well as both with DAPI (nuclear stain) and Sytox orange. No NET formation was observed when PA+Sias was incubated with neutrophils (A-C). In contrast, NETs formation was visualised after incubating PA-Sias with neutrophils followed by staining with Sytox orange only (B) as well as DAPI and Sytox orange together (D). Reproduced with permission from Society for Leukocyte Biology, Bethesda, USA (J Leukoc Biol 2012; 91: 641-55).
DNAse a glycosylated polypeptide, catalyzes the degradation of phosphodiester linkages of single- and double-stranded DNA. To address the question concerning the formation of NETs for killing PA- Sias, neutrophils are treated with DNAse. As a result, although there is no sialic acids-siglec-9 interaction of PA-Sias with DNAse-treated neutrophils, NETs formation is inhibited29. This is clearly reflected in higher survivability of PA-Sias confirming direct involvement of NETs for killing of PA-Sias.
In several vertebrates, different pathogenic microorganisms like S. aureus, Shigella flexneri, Streptococcus pyogenes, Bacillus anthracis, Mycobacterium tuberculosis, Candida albicans, and L. amazonensis are effectively killed through NETs formation26. Cathepsin G deficient-mice are more prone to S. aureus infections111. Similarly, neutrophil-elastase deficient mice are more susceptible to various bacteria like E. coli, K. pneumoniae and many enterobacterial infections108112. Staphylococcus aureus expressing nuclease, S. pneumoniae expressing capsule and D-alanylated lipoteichoic acid show resistance towards NETs-mediated killing by neutrophils resulting in increased pathogenesis in vivo2627.
Siglec-sialic acid interaction during Leishmania infection
During its life cycle Leishmania encounters a hostile environment and an array of immune effector cells. In due course of evolution, Leishmania has evolved mechanisms to survive and subvert the host immune system. Our recent findings indicate that Leishmania promastigotes interact more with siglec-1 and siglec-5 than other siglecs, indicating a specific sialic acids-siglec interaction might exist during infection (our unpublished data). Moreover, differential sialic acids-siglec interaction is observed among virulent and avirulent strain of L. donovani with different macrophage cell lines (unpublished data).
We further try to investigate whether sialic acids-siglec interaction modulate the immune response during leishmania infection. Preliminary findings reveal the phosphatase-mediated deactivation of various signaling pathways is differentially regulated in presence or absence of sialic acids-siglec interaction (Fig. 5, unpublished data).
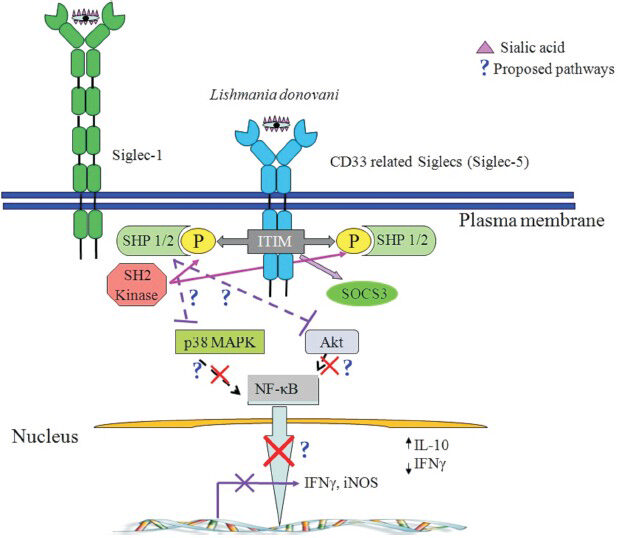
- Schematic diagram of the proposed pathway of CD33 siglec-mediated suppression of cellular function in Leishmania infection. On macrophage, after ligand (present on various pathogen surface) binding with CD33 related siglecs, immunoreceptor tyrosine-based inhibition motif (ITIM) present on the cytosolic portion of siglecs get activated by phosphorylation with SH2 family kinases (Lyn, Syk). Then Src homology 2 domain-containing phosphatases 1 and 2 (SHP1 and SHP2) bind with the ITIM and get activated. These activated SHPs further dephosphorylate various signaling molecules and suppress cellular activation. The target of SHPs might be p38 mitogen-activated protein kinases (p38 MAPK) or Akt or nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kβ) and ultimately downregulate the effector functions of the cell. In macrophages this siglec-mediated pathway might regulate the polarization of macrophage function towards anti-inflammatory type by reducing Th1 cytokines (IFNγ) and increasing Th2 cytokines (IL-10) along with downregulating inducible nitric oxide synthase (iNOS) gene expression for nitric oxide (NO) secretion.
Siglec-sialic acids interaction in Trypanosoma
Siglec-E, murine orthologue of human siglec-9 presents on neutrophils, macrophages and dendritic cells acts as a first line of defense against T.cruzi113. Pathogenic strain of T. cruzi exhibits enhanced binding with siglec-E-Fc chimera compared to non-pathogenic strain52. T. cruzi also contains the modified structure of sulphated glycan which is the ligand for siglec-E114. Siglec-E transfected CHO cells show increased association of parasites and rapid mobilization to the contact zone. However, control CHO cells and siglec-E transfected CHO cells exhibit same percentage of infection with T. cruzi, indicating that siglec-E is not an indispensable factor for infection53. T. cruzi interacts with tissue macrophages through sialic acids-siglec-1 dependent manner115. Dendritic cells reduce MHC (major histocompatibility complex) class-I antigen presentation in vitro116 and produce interleukin (IL)-12 which plays a major part against this infection113. Accordingly, there might be siglec-1-mediated association, phagocytosis of T. cruzi by macrophages and regulation of immune cells by ITIM bearing CD33-related siglecs.
Proposed mechanism of survival of PA in host
Taken together, we have demonstrated that pathogens adsorb sialic acids from the host serum and use these to inhibit C3-deposition28. Additionally, these also promote association with the innate immune cells through siglec-dependent recognition to persist within the host. Here, we have shown that a representative pathogen, PA, not only adsorbs sialic acids from host but also associates with neutrophils via the sialic acids-siglec-9 interaction. Due to such interaction, the neutrophils produce less ROS and elastase and thereby NETs formation is dramatically reduced29. Consequently, the neutrophil-mediated killing activity is impaired. Therefore, it may be envisaged that pathogens which are unable to synthesize sialic acids adsorb host sialic acids as a way to engage the inhibitory siglec-9 and thus subvert immunity (Fig. 6)29. This may be one of the general mechanisms by which other pathogens either synthesize or acquire sialic acids from host can establish infection by reducing the host innate immune response.
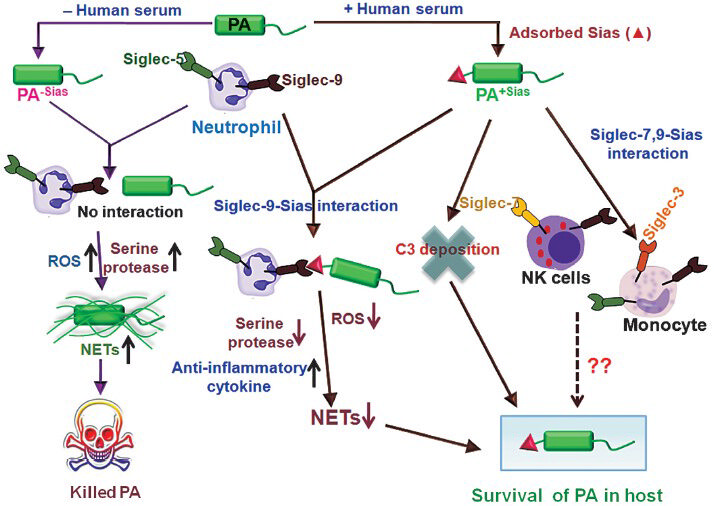
- Schematic diagram of proposed mechanism for survival of PA+Sias in hostile environment. Schematic representation of interaction of PA+Sias with immune cells through siglecs and their survival in host by dampening host innate immune response is shown. PA adsorbed sialic acids from host serum (PA+Sias). PA+Sias associated with immune cells (NK-cells, monocytes and neutrophils) through sialic acids-siglecs (siglec-7 and siglec-9, respectively) interaction. Neutrophils produced reduced amount of reactive oxygen species, serine proteases (elastase) and neutrophil extracellular traps (NETs) through siglec-9-sialic acids interaction with PA+Sias. At the same time, due to such interaction neutrophils enhanced anti-inflammatory cytokines production leading to survival of PA+Sias within host. Sialic acids on PA resist complement deposition directing survival of PA+Sias. On the other hand, PA cultured in sialic acids free medium (PA-Sias) exhibited no interaction with neutrophils. Therefore, neutrophils produced increased secretion of reactive oxygen species, serine proteases (elastase) followed by neutrophil extracellular traps (NETs) formation leading to effective clearance of PA. Reproduced with permission from Society for Leukocyte Biology, Bethesda, USA (J Leukoc Biol 2012; 91: 641-55).
Future perspective
In recent years, a great advancement has been made in the field of sialoglycobiology of human pathogens, still it is the tip of iceberg. The sialylation pattern and the mode of acquisition of sialic acids on several bacterial and viral families are yet to be explored. Although pathogens stimulate an array of inflammatory responses, the specific role of these sialylated molecules in evading the host defense remains to be fully characterized and appears to be a complex phenomenon. Even though siglec-mediated interaction has been established in a few pathogens, the role of siglecs in recognition and pathogenesis in other sialylated pathogens might open up a new avenue towards understanding the disease biology in near future. PA along with other important bacteria is becoming more and more antibiotic resistant day by day. Subsequently, the role of surface sialylation of pathogens and their siglec-mediated interaction with host immune cells might be helpful and possibly this information could be used as a potential therapeutic target as an alternative of antibiotics. The importance of this sialic acids-siglec interaction is not only restricted to the bacteria or in viruses, it is true even in parasitic infections such as leishmaniasis and trypanosomiasis. Such mechanisms used by the parasites to survive in the hostile environment are not well investigated till now, whereas more and more drug-resistant strains are arising. In this context, a detailed mechanistic approach is of utmost importance to explore sialic acids-siglec pathways for identifying future drug targets as sialic acids acquisition has become almost an indispensable factor for most of the human pathogens.
Acknowledgment
The work from the authors’ laboratory reported in this review was supported by the Council of Scientific and Industrial Research (CSIR), CSIR-Indian Institute of Chemical Biology, Department of Biotechnology (DBT) and Indian Council of Medical Research (ICMR), Government of India, New Delhi. The last author (CM) acknowledges the ICMR and the German Cancer Research Center for a mutual grant, and also DBT, Government of India, for J. C. Bose National Fellowship. The first two authors (BK, SR) acknowledge the CSIR, New Delhi for Senior Research Fellowship. Authors thank Prof. Richard D. Cummings of Emory University School of Medicine, Atlanta, GA, USA for providing the PA strain, Prof. Paul R. Crocker, College of Life Sciences, University of Dundee, UK, for providing human siglec-Fc chimera, CHO-siglec cells and anti-siglec antibodies; and Dr Alfredo Toreno, Servicio de Immunologia, Centro Nacional de Microbiologia, Instituto de salud Carlos III Majadahonda, Madrid, Spain and Dr R. Vlasak of Applied BioTechnology, Salzburg, Austria, for kind gift of anti-C3 antibody and 9-O-acetylesterase, respectively.
References
- Unified theory of bacterial sialometabolism: how and why bacteria metabolize host sialic acids. ISRN Microbiol. 2013;2013:8167-13.
- [Google Scholar]
- Sialic acids as regulators of molecular and cellular interactions. Curr Opin Struct Biol. 2009;5:507-14.
- [Google Scholar]
- O-acetylation of sialic acids- their detection, biological significance and alteration in diseases- a review. Trends Glycosci Glycotechnol. 2000;12:17-33.
- [Google Scholar]
- Investigation of 9-O-Acetylated sialoglycocongugates in childhood acute lymphoblastic leukaemia. Br J Hematol. 2000;110:801-12.
- [Google Scholar]
- Sialic acids in molecular and cellular interactions. Int Rev Cytol. 1997;175:137-240.
- [Google Scholar]
- Role of linkage specific 9-O-acetylated sialoglycoconjugates in activation of the alternate complement pathway on mammalian erythrocytes. Glycoconj J. 2000;17:887-93.
- [Google Scholar]
- Disease-associated glycosylated moleclar variants of human C-reactive protein activate complement-mediated hemolysis of erythrocytes in tuberculosis and Indian Visceral leishmaniasis. Glycoconj J. 2009;26:1151-69.
- [Google Scholar]
- Antibodies directed against O-acetylated sialoglycoconjugates accelerate complement activation in Leishmania donovani promastigotes. J Infect Dis. 2004;190:2010-9.
- [Google Scholar]
- Siglecs as targets for therapy in immune-cell-mediated disease. Trends Pharmacol Sci. 2009;30:240-8.
- [Google Scholar]
- Targeting Siglecs - A novel pharmacological strategy for immuno- and glycotherapy. Biochem Pharmacol. 2011;82:323-32.
- [Google Scholar]
- Siglec-9, a novel sialic acid binding member of the immunoglobulin superfamily expressed broadly on human blood leukocytes. J Biol Chem. 2000;275:22121-6.
- [Google Scholar]
- Siglec-8, A novel eosinophil specific member of the immunoglobulin superfamily. J Biol Chem. 2000;275:861-6.
- [Google Scholar]
- Identification of SAF-2, a novel siglec expressed on eosinophils, mast cells and basophils. J Allergy Clin Immunol. 2000;105:1093-100.
- [Google Scholar]
- Characterization of siglec-5, a novel glycoprotein expressed on myeloid cells related to CD33. Blood. 1998;92:2123-32.
- [Google Scholar]
- Identification and characterization of a novel Siglec, Siglec-7, expressed by human natural killer cells and monocytes. J Biol Chem. 1999;274:34089-95.
- [Google Scholar]
- Cloning, characterization, and phylogenetic analysis of siglec-9, a new member of the CD33-related group of siglecs. Evidence for co-evolution with sialic acid synthesis pathways. J Biol Chem. 2000;275:22127-35.
- [Google Scholar]
- Engagement of p75/AIRM1 or CD33 inhibits the proliferation of normal or leukemic myeloid cells. Proc Natl Acad Sci USA. 1999;96:15091-6.
- [Google Scholar]
- Initial and innate responses to viral infections - pattern setting in immunity or disease. Curr Opin Microbiol. 1999;2:374-81.
- [Google Scholar]
- Neutrophils: Cinderella of innate immune system. Int Immunopharmacol. 2010;10:1325-34.
- [Google Scholar]
- Isolation and phenotypic characterization of colonic macrophages. Clin Exp Immunol. 1998;112:205-15.
- [Google Scholar]
- Sialic acids acquired by Pseudomonas aeruginosa are involved in reduced complement deposition and siglec mediated host-cell recognition. FEBS Lett. 2010;584:555-61.
- [Google Scholar]
- Sialoglycoproteins adsorbed by Pseudomonas aeruginosa facilitate their survival by impeding neutrophil extracellular trap through siglec-9. J Leukoc Biol. 2012;91:641-55.
- [Google Scholar]
- Group B Streptococcal sialic acids interact with Siglecs (immunoglobulin-like lectins) on human leukocytes. J Bacteriol. 2007;189:1231-7.
- [Google Scholar]
- Genetic and biochemical modulation of sialic acid O-acetylation on group B Streptococcus: phenotypic and functional impact. Glycobiology. 2009;19:1204-13.
- [Google Scholar]
- Sialic acid: a preventable signal for pneumococcal biofilm formation, colonization, and invasion of the host. J Infect Dis. 2009;199:1497-505.
- [Google Scholar]
- Recognition of sialylated meningococcal lipopolysaccharide by siglecs expressed on myeloid cells leads to enhanced bacterial uptake. Mol Microbiol. 2003;49:1213-25.
- [Google Scholar]
- Requirement of NMB0065 for connecting assembly and export of sialic acid capsular polysaccharides in Neisseria meningitidis. Microbes Infect. 2010;12:476-87.
- [Google Scholar]
- Extracellular structure of polysialic acid explored by on cell solution NMR. Proc Natl Acad Sci USA. 2007;104:11557-61.
- [Google Scholar]
- Sialic acid-binding immunoglobulin-like lectin 7 mediates selective recognition of sialylated glycans expressed on Campylobacter jejuni lipooligosaccharides. Infect Immun. 2006;74:4133-41.
- [Google Scholar]
- Sialoadhesin promotes rapid proinflammatory and type I IFN responses to a sialylated pathogen, Campylobacter jejuni. J Immunol. 2012;189:2414-22.
- [Google Scholar]
- Visualization of sialic acid produced on bacterial cell surfaces by lectin staining. Microbes Environ. 2010;25:152-5.
- [Google Scholar]
- The lipooligosaccharides of Haemophilus ducreyi are highly sialylated. J Bacteriol. 1996;178:564-70.
- [Google Scholar]
- Chemo-enzymatic synthesis of the carbohydrate antigen N-glycolylneuraminic acid from glucose. Carbohydr Res. 2010;345:1225-9.
- [Google Scholar]
- Variable degree of alternative complement pathway-mediated hemolysis in Indian visceral leishmaniasis induced by differential expression of 9-O-acetylated sialoglycans. J Infect Dis. 2004;189:1257-64.
- [Google Scholar]
- Sialoglycans in protozoal diseases: their detection, modes of acquisition and emerging biological roles. Glycoconj J. 2004;20:199-206.
- [Google Scholar]
- Identification of sialic acids on Leishmania donovani amastigotes. Biol Chem. 2004;385:59-66.
- [Google Scholar]
- O-acetyl sialic acids in parasitic diseases. In: Yerema KJ, ed. Hand book of carbohydrate engineering. USA: Taylor and Francis Group; 2005. p. :71-98.
- [Google Scholar]
- Targeting glycoproteins or glycolipids and their metabolic pathways for anti-parasite therapy. Adv Exp Med Biol. 2008;625:87-102.
- [Google Scholar]
- A perspective on the emergence of sialic acids as potent determinants affecting Leishmania biology. Mol Biol Int. 2011;2011:532106.
- [Google Scholar]
- Identification and characterization of adsorbed serum sialoglycans on Leishmania donovani promastigotes. Glycobiology. 2003;13:351-61.
- [Google Scholar]
- 9-O-acetylated sialic acids enhance entry of virulent Leishmania donovani promastigotes into macrophages. Parasitology. 2009;136:159-73.
- [Google Scholar]
- Sialic acids in different Leishmania sp., its correlation with nitric oxide resistance and host responses. Glycobiology. 2010;20:553-66.
- [Google Scholar]
- Increased association of Trypanosoma cruzi with sialoadhesin positive mice macrophages. Parasitol Res. 2005;97:380-5.
- [Google Scholar]
- Sialylated ligands on pathogenic Trypanosoma cruzi interact with Siglec-E (sialic acid-binding Ig-like lectin-E) Cell Microbiol. 2009;11:1600-11.
- [Google Scholar]
- Molecular interaction of Siglecs (sialic acid-binding Ig-like lectins) with sialylated ligands on Trypanosoma cruzi. Eur J Cell Biol. 2010;89:113-6.
- [Google Scholar]
- Siglecs facilitate HIV-1 infection of macrophages through adhesion with viral sialic acids. PLoS One. 2011;6:e24559.
- [Google Scholar]
- Porcine arterivirus infection of alveolar macrophages is mediated by sialic acid on the virus. J Virol. 2004;78:8094-101.
- [Google Scholar]
- The M/GP(5) glycoprotein complex of porcine reproductive and respiratory syndrome virus binds the sialoadhesin receptor in a sialic acid-dependent manner. PLoS Pathog. 2010;6:e1000730.
- [Google Scholar]
- Identification and analysis of O-acetylated glycoproteins. Methods Mol Biol. 2013;981:57-93.
- [Google Scholar]
- The elderberry (Sambucus nigra L.) bark lectin recognizes the Neu5Ac(alpha 2-6)Gal/GalNAc sequence. J Biol Chem. 1987;262:1596-601.
- [Google Scholar]
- The immobilized leukoagglutinin from the seeds of Maackia amurensis binds with high affinity to complex-type Asn-linked oligosaccharides containing terminal sialic acid-linked alpha-2,3 to penultimate galactose residues. J Biol Chem. 1988;263:4576-85.
- [Google Scholar]
- Lectin structure-activity: the story is never over. J Agric Food Chem. 2002;50:6583-5.
- [Google Scholar]
- The specificity of the binding site of Achatinin-H, a sialic-acid binding lectin from Achatina fulica. Carbohydr Res. 1995;268:115-25.
- [Google Scholar]
- Identification of antibodies directed against O-acetylated sialic acids in visceral leishmaniasis: its diagnostic and prognostic role. Glycoconj J. 1998;15:1141-7.
- [Google Scholar]
- Isolation and properties of two sialate-O-acetylesterases from horse liver with 4- and 9-O-acetyl specificities. Glycoconj J. 2008;25:625-32.
- [Google Scholar]
- Rapid diagnosis of Indian visceral leishmaniasis using AchatininH, a 9-O-acetylated sialic acid binding lectin. Am J Trop Med Hyg. 1998;58:551-4.
- [Google Scholar]
- Functions and Biosynthesis of O-acetylated sialic acids. Top Curr Chem 2012 In press
- [Google Scholar]
- Regulation of O-acetylation of sialic acids by sialate-O-acetyltransferase and sialate-O-acetylesterase activities in childhood acute lymphoblastic leukemia. Glycobiology. 2012;22:70-83.
- [Google Scholar]
- Trypanosoma cruzi surface mucins: host-dependent coat diversity. Nat Rev Microbiol. 2006;4:229-36.
- [Google Scholar]
- To sialylate, or not to sialylate: that is the question. Trends Microbiol. 2002;10:254-7.
- [Google Scholar]
- Preliminary 1H NMR investigation of sialic acid transfer by the trans-sialidase from Trypanosoma cruzi. Bioorg Med Chem Lett. 2000;10:2791-4.
- [Google Scholar]
- Role of sialic acid in the resistance of Trypanosoma cruzi trypomastigotes to complement. J Immunol. 1994;153:3141-7.
- [Google Scholar]
- DNA markers define two major phylogenetic lineages of Trypanosoma cruzi. Mol Biochem Parasitol. 1996;83:141-52.
- [Google Scholar]
- Differential expression of a virulence factor, the trans-sialidase, by the main Trypanosoma cruzi phylogenetic lineages. J Infect Dis. 2004;189:2250-9.
- [Google Scholar]
- Diversity of microbial sialic acid metabolism. Microbiol Mol Biol Rev. 2004;68:132-53.
- [Google Scholar]
- Analysis of complement C3 deposition and degradation on Klebsiella pneumoniae. Infect Immun. 1996;64:4726-32.
- [Google Scholar]
- Functional comparison of the binding of factor H short consensus repeat 6 (SCR 6) to factor H binding protein from Neisseria meningitidis and the binding of factor H SCR 18 to 20 to Neisseria gonorrhoeae porin. Infect Immun. 2009;77:2094-103.
- [Google Scholar]
- Capsular sialic acid limits C5a production on type III group B streptococci. Infect Immun. 1999;67:1866-70.
- [Google Scholar]
- Development of an assay for quantification of linkage-specific O-acetylated sialoglycans on erythrocytes; its application in Indian visceral leishmnaiasis. J Immunol Methods. 2002;270:1-10.
- [Google Scholar]
- Identification of 9-O-acetylated sialoglycans on peripheral blood mononuclear cells in Indian visceral leishmaniasis. Glycoconj J. 2004;20:531-6.
- [Google Scholar]
- 9-O-acetylated sialoglycoproteins: Important immunomodulators in Indian visceral leishmaniasis. Clin Vaccine Immunol. 2009;18:889-98.
- [Google Scholar]
- Detection and characterization of a sialoglycosylated bacterial ABC-type phosphate transporter protein from patients with visceral leishmaniasis. Glycoconj J. 2009;26:675-89.
- [Google Scholar]
- Glycosylation of erythrocyte spectrin and its modification in visceral leishmaniasis. PLoS One. 2011;6:e28169.
- [Google Scholar]
- Sialoglycosylation of RBC in visceral leishmaniasis leads to enhanced oxidative stress, calpain-induced fragmentation of spectrin and hemolysis. PLoS One. 2012;7:e42361.
- [Google Scholar]
- The membrane-proximal immunoreceptor tyrosine based inhibitory motif is critical for the inhibitory signaling mediated by Siglecs-7 and-9, CD33-related Siglecs expressed on human monocytes and NK cells. J Immunol. 2004;173:6841-9.
- [Google Scholar]
- Discovery of Siglec-14, a novel sialic acid receptor undergoing concerted evolution with Siglec-5 in primates. FASEB J. 2006;20:1964-73.
- [Google Scholar]
- Sampling and signaling in plasmacytoid dendritic cells: the potential roles of Siglec-H. Trends Immunol. 2006;27:255-60.
- [Google Scholar]
- Masking of CD22 by cis ligands does not prevent redistribution of CD22 to sites of cell contact. Proc Natl Acad Sci USA. 2004;101:6104-9.
- [Google Scholar]
- Characterization of CD33 as a new member of the sialoadhesin family of cellular interaction molecules. Blood. 1995;85:2005-12.
- [Google Scholar]
- Properties and distribution of a lectin-like hemagglutinin differentially expressed by murine stromal tissue macrophages. J Exp Med. 1986;164:1862-75.
- [Google Scholar]
- Sialoadhesin binds preferentially to cells of the granulocytic lineage. J Clin Invest. 1995;95:635-43.
- [Google Scholar]
- Masking and unmasking of the sialic acid-binding lectin activity of CD22 (Siglec-2) on B lymphocytes. Proc Natl Acad Sci USA. 1998;95:7469-74.
- [Google Scholar]
- A novel subset of murine B cells that expresses unmasked forms of CD22 is enriched in the bone marrow: implications for B-cell homing to the bone marrow. Immunology. 2000;101:342-7.
- [Google Scholar]
- Cryptic sialic acid binding lectins on human blood leukocytes can be unmasked by sialidase treatment or cellular activation. Glycobiology. 1999;9:1225-34.
- [Google Scholar]
- High-affinity ligand probes of CD22 overcome the threshold set by cis ligands to allow for binding, endocytosis, and killing of B cells. J Immunol. 2006;177:2994-3003.
- [Google Scholar]
- Ganglioside GD3 expression on target cells can modulate NK cell cytotoxicity via siglec-7-dependent and-independent mechanisms. Eur J Immunol. 2003;33:1642-8.
- [Google Scholar]
- Probing the cis interactions of the inhibitory receptor Siglec-7 with α2,8-disialylated ligands on natural killer cells and other leukocytes using glycan-specific antibodies and by analysis of α2,8-sialyltransferase gene expression. J Leukoc Biol. 2006;80:787-6.
- [Google Scholar]
- Involvement of sialoadhesin in entry of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages. J Virol. 2003;77:8207-15.
- [Google Scholar]
- Molecular mimicry of host sialylated glycans allows a bacterial pathogen to engage neutrophil siglec-9 and dampen the innate immune response. Blood. 2009;113:3333-6.
- [Google Scholar]
- Group B Streptococcus suppression of phagocyte functions by protein-mediated engagement of human siglec-5. J Exp Med. 2009;206:1691-9.
- [Google Scholar]
- Characterization of the specific interaction between sialoadhesin and sialylated Campylobacter jejuni lipooligosaccharides. Infect Immun. 2010;78:3237-46.
- [Google Scholar]
- Reactive oxygen and nitrogen species as signaling molecules regulating neutrophil function. Free Radic Biol Med. 2007;42:153-64.
- [Google Scholar]
- Oxidative innate immune defenses by Nox/Duox family NADPH oxidases. Contrib Microbiol. 2008;15:164-87.
- [Google Scholar]
- Neutrophil-mediated oxidative burst and host defense are controlled by a Vav-PLCgamma2 signaling axis in mice. J Clin Invest. 2007;117:3445-52.
- [Google Scholar]
- Clinical Immunology Review Series: An approach to the patient with recurrent superficial abscesses. Clin Exp Immunol. 2008;152:397-405.
- [Google Scholar]
- Neutrophil Elastase: Path Clearer, Pathogen Killer, or Just Pathologic? Am J Respir Cell Mol Biol. 2002;26:266-8.
- [Google Scholar]
- Neutrophil elastases targets virulence factors of enterobacteria. Nature. 2002;417:91-4.
- [Google Scholar]
- The periplasmic serine protease inhibitor ecotin protects bacteria against neutrophil elastase. Biochem J. 2004;379:107-18.
- [Google Scholar]
- Killing activity of neutrophils is mediated through activation of proteases by K+ flux. Nature. 2002;416:291-7.
- [Google Scholar]
- Mice lacking neutrophil elastases reveal impaired host defense against gram negative bacterial sepsis. Nat Med. 1997;4:615-8.
- [Google Scholar]
- NK cells contribute to the control of Trypanosoma cruzi infection by killing free parasites by perforin-independent mechanisms. Infect Immun. 2004;72:6817-25.
- [Google Scholar]
- Sulfated lipids represent common antigens on the surface of Trypanosoma cruzi and mammalian tissues. Mol Biochem Parasitol. 1988;30:113-21.
- [Google Scholar]
- Trypanosoma cruzi down-regulates lipopolysaccharide-induced MHC class I on human dendritic cells and impairs antigen presentation to specific CD8+ T lymphocytes. Int Immunol. 2002;14:1135-44.
- [Google Scholar]
- Visceral leishmaniasis: elimination with existing interventions. Lancet Infect Dis. 2011;11:322-5.
- [Google Scholar]




