Translate this page into:
Role of hepatitis B virus genotype D & its mutants in occult hepatitis B infection
Reprint requests: Dr Subrat Kumar Panda, Department of Pathology, All India Institute of Medical Sciences Ansari Nagar, New Delhi 110 029, India e-mail: subrat@gmail.com
-
Received: ,
This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-Share Alike 3.0 Unported, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This article was originally published by Medknow Publications & Media Pvt Ltd and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Background & objectives:
Non-detection of hepatitis B virus (HBV) envelope protein (hepatitis B surface antigen, HBsAg) in a chronically HBV infected individual has been described as occult infection. One possible reason for this phenotype is alteration in large (L-HBsAg) to small (S-HBsAg) envelope protein ratio associated with reduced or non secretion of HBsAg. This results in quantitative levels of serum HBsAg below the detection limit of enzyme immunoassays. Genotype D of HBV has a characteristic 33 nucleotide (nt) deletion upstream of the pre-S2/S promoter. This deletion may reduce HBsAg secretion in occult infection patients infected with genotype D HBV. Additional deletions in the pre-S2/S promoter may further aggravate reduced HBsAg secretion in patients infected with genotype D HBV. Thus, the aim of the present study was to determine the role of genotype D specific 33nt deletion and additional pre-S2/S promoter deletions in causing reduced or no secretion of HBsAg, in occult infection. Since these deletions overlap virus polymerase, their effect on virus replication was also investigated.
Methods:
We examined the in vitro expression of HBsAg, ratio of cure and ‘e’ antigen (HBcAg/HBeAg), their secretion and virus replication, using overlength 1.3 mer/1.86 mer genotype A replicons, and genotype D replicons with and without additional pre-S2/S promoter deletions from cases of occult infection.
Results:
Genotype D replicon showed a decrease in HBsAg secretion compared to the wild-type genotype A. Genotype D replicons carrying additional pre-S2/S promoter deletions, showed further reduction in HBsAg secretion, demonstrated presence of intracellular HBcAg/HBeAg, virus replication intermediates and ‘e’ antigen secretion.
Interpretation & conclusions:
The characteristic 33 nt deletion of genotype D HBV reduces HBsAg secretion. Additional pre-S2/S promoter deletions may further diminish HBsAg secretion, leading to occult infection. Pre-S2/S promoter deletions do not affect HBV replication.
Keywords
Genotypes
HBV
occult
pre-S2/S
promoter mutants
surface antigen (HBsAg)

Hepatitis B virus (HBV) infection is associated with acute liver disease, including liver failure and chronicity that can lead to cirrhosis and liver cancer1. Currently eight HBV genotypes (A-H) have been described which diverge by more than 8 per cent in their nucleotide sequences2. HBV genotype D has a characteristic deletion of 33 nucleotides (33 nt) upstream of the pre-S2/S promoter, within the pre-S1 open reading frame (ORF)2.
Occult hepatitis B virus infection is defined as “the presence of HBV DNA in the liver (with detectable or undetectable HBV DNA in serum) in individuals testing HBsAg negative by currently available assays”34. Earlier, naturally occurring deletions in the pre-S2/S promoter region were observed in several cases of occult HBV infection56, chronic HBV infection7, and patients with progressive liver diseases8. In a subsequent study, it was demonstrated that these deletions can cause altered surface protein expression, and an increased large-HBsAg (L-HBsAg) to major/small-HBsAg (S-HBsAg) ratio leading to reduced HBsAg secretion9. However, in this study9 the characteristic genotype D specific 33 nt deletion was observed in the HBV surface promoter surface gene fusion constructs used. In another study, least quantities of secreted HBsAg were observed in HBV genotype D10. Genotype D has been found to be associated with occult hepatitis B infection1112. In a recent study, diagnostic PCR for the pre-S1 coding region of HBV isolates obtained from 57 occult infection patients, (who were HBsAg negative, anti-HBc and HBV DNA positive) revealed that all HBV isolates showed the characteristic 33 nt deletion of genotype D13. The above studies point to a possible role of the genotype D specific deletion in causing occult hepatitis B infection. Thus in the current study, we analysed the role of the genotype D specific 33 nt deletion in reducing HBsAg secretion. Additional pre-S2/S promoter deletions in such cases were analysed for further aggravating this phenotype. Since these deleted regions overlap virus polymerase, the replication competence of mutants carrying these deletions was also investigated.
Material & Methods
The study was carried out in the department of Pathology, All India Institute of Medical Sciences (AIIMS), New Delhi, India from April 2007 to November 2011.
The study was carried out with HBV isolates derived from occult hepatitis B infection patients [313(S5), 761(S8), 821(S9)] selected from a previous study5. Overlength 1.86mer/1.3mer replicons of these isolates were made which consisted of the pre-S1, pre-S2/S promoter region of these isolates (consisting of the 33 nt genotype D specific deletion with or without additional pre-S2/S promoter deletions) in a wild-type genotype A background, to prevent variations in HBV replicons other than the specific deletions under investigation. These replicons were analysed for in vitro expression and secretion of HBsAg to determine the role of these deletions in causing reduced HBsAg secretion in occult hepatitis B infection. Two of the 1.86mer replicons carrying pre-S2/S promoter deletions, in addition to the genotype D specific 33 nt deletion were checked for their ability to affect virus replication.
Diagnosis, serological characteristics and immuno-histochemisty results of liver biopsies of selected patients whose HBV isolates were used in the current study are listed elsewhere5. All three patients were negative for serum HBsAg and positive for serum HBV DNA5.
Construction of overlength 1.86 mer/1.3 mer genotype A and D replicons: The numbering of the circular HBV genome (GenBank Accession no. X02763, length 3,221 base pairs) was used with the numbering at the EcoR1 site as 3219G3220A3221A1T2T3T2,5. Overlength 1.86mer wild type HBV DNA replicon (GenBank Accession no. X02763, Genotype A, subtype adw2) was produced in pRLnullΔ(77-1246) (i.e. pRLnull with the T7 promoter and Renilla luciferase gene deleted) vector and served as the control. This construct was designated as pRLnullΔ(77-1246)WT-1.86. Both sets of pre-S1, pre-S2/S promoters in the wild type genotype A replicon were replaced with pre-S1 and pre-S2/S promoter sequences (nt 2600- nt 3221/1- nt 3) from HBV clones 821.1, 313.1, 761.1, (Genbank accession nos: AY161161, AY161147, AY161159) derived from occult infection patients 821, 313, 761, respectively. This was done to prevent variations in the HBV replicons other than the mutations being investigated. The constructs pRLnullΔ(77-1246)WT-1.86, pRLnullΔ(77-1246)821.1-1.86 pRLnullΔ(77-1246)313.1-1.86 and pRLnullΔ(77-1246)761.1-1.86 will henceforth be referred to as WT-1.86, 821.1-1.86, 313.1-1.86 and 761.1-1.86. The constructs were sequence confirmed using big dye terminator v3.1cycle sequencing kit (Applied Biosystems, USA) and ABI prism 310 genetic analyzer (USA).
Overlength 1.3 mer constructs were constructed as follows: pRLnull(Δ77-1246)WTSPGE, coding for HBV Large, Middle, Small-HBsAg promoters, genes and enhancers I,II9 was digested with BglII (nt 2429) and EcoRV (nt 1042). The BglII 5’ overhang was filled with T4 DNA polymerase and ligated to the blunt end produced by EcoRV resulting in the vector backbone pRLnullΔ(77-1246)WTSPGEΔ (2429-1042). The vector backbone consisted of HBV genome sequences from nucleotides 1042-1988 (~0.3mer). Overlength 1.3mer wild-type genotype A construct was produced by inserting the 3221 base pair (1.0mer) SacII fragment from pRLnullΔ (77-1246)WT-1.86 into SacII digested pRLnullΔ(77-1246)WTSPGEΔ(2429-1042). Similar insertions were carried out for producing overlength genotype D, 821.1-1.3mer, 313.1-1.3mer, 761.1-1.3mer constructs. The constructs pRLnullΔ(77-1246)WT-1.3, pRLnullΔ(77-1246)821.1-1.3, pRLnullΔ(77-1246) 313.1-1.3 and pRLnullΔ(77-1246)761.1-1.3 will henceforth be referred to as WT-1.3, 821.1-1.3, 313.1-1.3 and 761.1-1.3.
HBV has a circular genome with overlapping open reading frames. However, studies for subtype adw2, which is the wild-type used in the present study have been carried out using linear overlength (1.3/1.5mer) replication competent constructs consisting of two sets of enhancer, (enhancers I and II) one at the 5’ end and another at the 3’ end of the linear construct1415. The enhancers at the 3’end of the linear construct are required for pre-S1 and pre-S2 promoter function1617. Enhancer I at the 5’end of the construct is required for normal expression of all the viral transcripts18. However, since the 1.3mer constructs carry two sets of enhancers for a single set of pre-S2/S promoters, these have been observed to produce larger amounts of HBsAg compared to constructs with a single set of enhancers16. Over-length 1.86mer HBV constructs were used because we hypothesized that these would be maintaining the large versus small envelope protein ratio (L/S) since for both sets of enhancers there are two sets of pre-S2/S promoters (Fig. 1A). Thus we wanted to determine whether the L/S ratio plays a strict role in regulating HBsAg secretion, or whether the total amounts of surface proteins affect HBsAg synthesis, and secretion. Further, in the overlength 1.86mer construct a single viral polymerase open reading frame is present (Fig. 1A). Hence in the current study the viral replication intermediates are produced by a single viral polymerase.

-
(A) Schematic representation (not drawn to scale) of overlength 1.86mer/1.3mer constructs of HBV. The over-length 1.86mer construct begins at nucleotide 2429 while the 1.3mer construct begins at nucleotide 1045. Block arrows indicate HBV promoters. Grey rectangularboxes depict open reading frames and enhancers of HBV. (B) Schematic representation (not drawn to scale) of deletions of the various occult HBV infection isolates. Genotype D HBV isolates 821.1, 313.1, 761.1 show deletions of 33 nucleotides (Δnt 2859-2878, 2882-2894) upstream of the pre-S2/S promoter in the pre-S1ORF. HBV isolate 313.1 carries pre-S2/S promoter deletions (Δnt 3026-3208). HBV isolate 761.1 carries pre-S2/S promoter deletions (Δnt 3039-3095).
Construct pRLnullΔ (77-1246)WTSPGE coding for HBV Large, Middle, Small-HBsAg promoters, genes and enhancers I,II9 incapable of replication was used as negative control in some experiments. The negative control plasmid vector was used so that DNA detected by the HBV probe in Southern blot is specific for HBV DNA sequences produced by virus replication.
Maintenance of Huh7 cells and transfections: Huh7 human hepatoma cell lines were cultured in maintenance medium9. Cells were transfected with overlength 1.86mer/1.3mer, genotype A and D replicons, using lipofectamine 2000 (Invitrogen, USA) according to manufacturer's instructions. pcDNA3Fluc (pcDNA3 Firefly luciferase) was co-transfected with the replicons for normalizing transfection efficiency and cell lysis detection. In all cases, the culture supernatants and transfected cells were processed for HBsAg ELISA. In case of the 313-1.86mer, 761.1-1.86mer constructs, cells were processed for HBeAg ELISA (for ‘e’ antigen), indirect immunofluorescence for HbcAg (for core antigen), Southern hybridization and real time PCR.
Detection of extracellular and intracellular HBsAg, HBeAg by ELISA: The culture supernatants were aspirated and adherent cells were washed three times, trypsinized and harvested. Equal number of viable adherent cells were pelleted and lysed separately with radio-immunoprecipitation assay (RIPA) buffer [50 mM Tris pH 7.4, 140mm NaCl, 0.5% Triton X-100, 0.05% w/v sodium dodecyl sulphate (SDS), 1 % w/v sodium deoxycholate, 5mM iodoacetamide and 2mM phenylmethylsulphonyl fluoride] and HBeAg lysis buffer (50mM Tris pH 8.0, 1% v/v NP-40, 100 mM NaCl and 0.1% v/v β-mercaptoethanol) for 20 min at 4oC. The lysates were clarified by centrifugation and the supernatants were used as the clarified cellular lysate. Culture supernatant and clarified cellular lysate were processed for detection of HBsAg at 48 h post-transfection using the Monolisa HBsAg ULTRA kit (Biorad, USA) and HBeAg, at 72 h post-transfection using the Monolisa HBe Ag-Ab PLUS kit (Biorad, USA) as per the manufacturers’ instructions. The ELISA readings were taken in the Sunrise TECAN ELISA plate reader, USA [with measurement range 0.0-4.0 optical density (OD) units, and linearity between 0.0-3.0 OD units]. All cell lysates and supernatants were diluted appropriately (supernatants were diluted 1:50 to 1:100 and lysates were diluted 1:10 to 1:20) so that ELISA OD readings were completely within the linear range, i.e. between 0.0 and 1.0.
Intracellular expression pattern of hepatitis B core antigen (HBcAg) by indirect immunofluorescence: Huh7 cells on a 22mm glass cover slip in a 30mm Petri dish (Nunc, USA) were transfected with each of the constructs 313.1-1.86, 761.1-1.86. Seventy two hours post transfection the cells were processed for HBcAg immunofluorescence as mentioned previously9. The primary antibody used was 1:100 dilution of 1mg of rabbit polyclonal anti-HBcAg ml-1 (HBVcAg-Ab-1, Thermo scientific, CA, USA).
Isolation of intracellular DNA followed by detection of HBV replication intermediates by Southern blot and quantitation by real time PCR: Seventy two hours post-transfection, the adherent cells were washed, trypsinized, harvested and suspended in 1ml 1X PBS. A 100 μl aliquot was processed for luciferase assay (Promega, USA). The other 900 μl aliquot was pelleted and lysed in 400 μl HBeAg lysis buffer for 20 min at 4°C, followed by centrifugation. The supernatants were used as the clarified cellular lysate. The cell lysates were processed as follows: the cell lysate was prepared with HBeAg lysis buffer, as described above, to release intracellular HBV nucleocapsids. The cellular lysate (100 μl) was treated with 300 μg DNAse 1/ml (Bangalore Genei, Bengaluru, India) to digest the plasmid DNA for 1 h at 37°C, twice. RNAse A (100 μg/ml Fermentas, Canada) was also added to digest the cellular RNA. This was followed by Proteinase K (Bangalore Genei, Bengaluru, India) digestion of virus capsids with its buffer (20mM EDTA, 0.5% w/v SDS, 2mM CaCl2) for 2 h at 55°C at a concentration of 600 μg/ml. The released virus DNA was extracted with Tris-EDTA saturated phenol, pH 8.0 (Sigma, USA) and ethanol precipitated with 7.5M ammonium acetate. The DNA was pelleted, washed, and dissolved in 30 μl water. A 5 μl aliquot of the DNA was processed for Taqman based quantitative real time PCR19 using the ABI prism 7700 sequence detection system, USA.
Southern hybridization20 was done with 25 μl aliquot of the DNA from mock transfected cells and appropriate volumes of the DNA from cells transfected with various plasmid constructs after normalizing for transfection efficiency and adjusting for the relative number of molecules transfected. Probe was prepared from a linear wild type 3.2 kb HBV DNA template with α-32P (BRIT, Mumbai, India), by random priming using a deca-labelling kit (Fermentas, Canada).
Statistical analysis: Extracellular and intracellular HBsAg was calculated in genotype D replicons, relative to wild-type genotype A. Unpaired t-test was used to calculate the difference in means (graph pad, PRISM software), USA. Percentage HBsAg secretion was calculated from the ratio of secreted versus total HBsAg in all the constructs relative to wild-type genotype A. In real time PCR, intracellular (IC) HBV DNA copy number (for mutant) over background [pRlnullΔ(77-1246)WTSPGE] normalized for transfection efficiency was calculated as follows:
HBV DNA copy number fold over background normalized for tranfection efficiency =
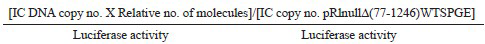
The relative number of molecules of DNA transfected was taken into consideration since the negative control plasmid pRlnullΔ (77-1246) WTSPGE was smaller in size (~4.9 kb) compared to the 1.86mer plasmids (~8.0 kb). Hence, for the same amount (in μg) of DNA transfected, the negative control would have greater number of molecules than the 1.86mer construct.
Results
Replicons 821.1, 313.1 and 761.1 carried the genotype D specific 33nt deletion from nt 2859-2878, 2882-2894 (Fig. 1B) upstream of the pre-S2/S promoter, in the pre-S1 ORF. In the polymerase open reading frame these will correspond to in-frame deletions from amino acids 185-195. Pre-S2/S promoter deletions in HBV isolates 313.1 and 761.1 have been described earlier9 [Figure1]B. These deletions overlap the polymerase open reading frame, correspond to in-frame deletions from amino acids 240-300 in the virus polymerase spacer region for HBV isolate 313.1 and amino acids 243-261 for HBV isolate 761.1.
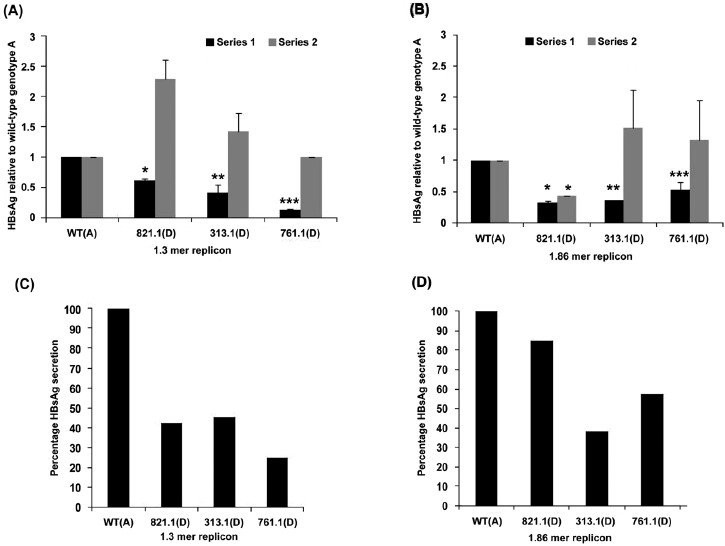
HBsAg secretion and expression in genotype D replicons relative to wild-type genotype A: Figs 2A and 2B depict extracellular (black bars) and intracellular (gray bars) HBsAg, in the over-length 1.3mer and 1.86mer genotype D replicons, respectively, relative to the wild-type genotype A. The luciferase readings in the culture supernatants were 0.1-0.8 per cent of the readings in cell lysates, indicating that no or minimal cell lysis had occurred. Thus the extracellular HBsAg represented secreted HBsAg. Extracellular HBsAg for wild-type genotype A was taken to be 1.0. At 48 h post-transfection, extracellular HBsAg in the genotype D replicon, 821.1-1.3mer, was significantly reduced (0.615 ± 0.04) (Fig. 2A). Further reduction in extracellular HBsAg was observed for the 313.1-1.3mer (0.42 ± 0.13) and 761.1-1.3mer (0.145 ± 0.01) (P<0.01) replicons which carried additional pre-S2/S promoter deletions (Fig. 2A). Differences in intracellular HBsAg for all the 1.3mer genotype D replicons relative to the wild-type genotype A, and relative to each other were insignificant (Fig. 2A). These results were recapitulated when the per cent HBsAg secretion was calculated (which is the amount of extracellular HBsAg divided by the total amount of HBsAg synthesized for the 1.3mer replicons) (Fig. 2C). Extracellular HBsAg in case of 821.1-1.86mer and 313.1-1.86mer replicons was significantly less (0.32 ± 0.03 and 0.36, respectively) than that of wild-type genotype A (1.0) (Fig. 2B). Replicon 761.1-1.86mer demonstrated less extracellular HBsAg (0.533+0.12, P=0.07, Fig. 2B) relative to wild-type genotype A but greater than replicons 821.1-1.86mer and 313.1-1.86mer (Fig. 2B). However, the differences in extracellular HBsAg between 821.1-1.86mer, 313.1-1.86mer, and 761.1-1.86mer replicons was not significant. No significant difference was observed in intracellular HBsAg between the wild-type genotype A and 313.1-1.86 and 761.1-1.86 replicons, and 821.1-1.86 and 313.1-1.86 and 761.1-1.86 replicons (Fig. 2B). However, the 821.1-1.86 genotype D replicon showed significantly less intracellular HBsAg relative to the wild-type genotype A (Fig. 2B). The same result in terms of per cent HBsAg secretion demonstrated a reduction in per cent HBsAg secretion for 821.1-1.86mer in comparison to genotype A. Further reduction in per cent HBsAg secretion was observed for the 313.1-1.86 mer and 761.1-1.86 mer replicons. This is because the mean levels of intracellular HBsAg for 313.1-1.86mer and 761.1-1.86 mer was greater than that for 821.1-1.86 mer. Thus it can be concluded that the characteristic 33 nucleotide deletion of genotype D has a role in reducing HBsAg secretion. Additional pre-S2/S promoter deletions may further diminish HBsAg secretion.
-
(A) Extracellular HBsAg (series 1, black bars) and intracellular HBsAg (series 2, gray bars) in Huh7 cells transfected with the wild-type (WT)-1.3 (genotype A), 821.1-1.3 (genotype D), 313.1-1.3 (genotype D), and 761.1-1.3 (genotype D) constructs at 48 h post-transfection. P values are relative to the wild-type genotype A (P*<0.001, **< 0.05, ***<0.005). (B) Extracellular HBsAg (series 1, black bars) and intracellular HBsAg (series 2, gray bars) in Huh7 cells transfected with the WT-1.86 (genotype A), 821.1-1.86 (genotype D), 313.1-1.86 (genotype D), and 761.1-1.86 (genotype D) constructs at 48 h post-transfection. P values are relative to the wild-type genotype A. (P*<0.005, **<0.0001, ***<0.1, (n=3). (C) Percentage HBsAg secretion in Huh7 cells transfected with the WT-1.3 (genotype A), 821.1-1.3 (genotype D), 313.1-1.3 (genotype D), and 761.1-1.3 (genotype D) constructs at 48 h post-transfection. (D) Percentage HBsAg secretion in Huh7 cells transfected with the WT-1.86 (genotype A), 821.1-1.86 (genotype D), 313.1-1.86 (genotype D), and 761.1-1.86 (genotype D) constructs at 48 h post-transfection.
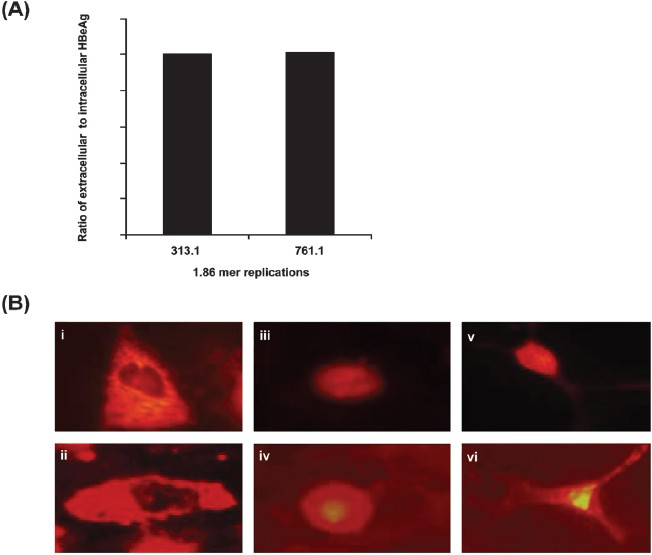
Pre-core/core antigen production and processing in the overlength 313.1mer and 761.1mer replicons: HBeAg production and secretion in the overlength 313.1-1.86mer and 761.1-1.86mer replicons were observed (Fig. 3A). Cells transfected with the 313.1-1.86mer and 761.1-1.86mer (Fig. 3B) replicons showed cells with only cytoplasmic HBcAg staining (Fig. 3B-i, ii), only nuclear HBcAg staining (Fig. 3B-iii,iv) and both cytoplasmic and nuclear HBcAg staining (Fig. 3B-v, vi). These results indicate that expression of ‘e’/core antigen and secretion of ‘e’ antigen occurs in these replicons.
-
(A) Ratio of extracellular to intracellular HBeAg in Huh7 cells transfected with the 313.1-1.86 and 761.1-1.86 constructs at 48 h post-transfection. (B) Intracellular expression pattern of HBcAg by indirect immunofluorescence in Huh7 cells transfected with the 313.1-1.86 and 761.1-1.86 constructs. The Figure depicts cytoplasmic staining (i, ii), nuclear staining (iii, iv) and combined cytoplasmic and nuclear staining (v, vi) of Huh7 cells transfected with the mutant 313.1-1.86 (i,iii,v) and mutant 761.1-1.86 constructs (ii, iv, vi), respectively for HBcAg.
Replication of 313.1-1.86mer and 761.1-1.86mer replicons: The mutant 313.1-1.86mer and 761.1-1.86mer replicons showed an approximately 3.2 kb intracellular double stranded linear virus DNA with a smear of virus replication intermediates by Southern blot. Relaxed circular virus DNA migrating slightly slower than linear double stranded DNA was also observed in case of the 761.1-1.86mer replicon (Fig. 4A). Replicons 313.1-1.86 and 761.1-1.86 showed approximately thousand fold greater DNA compared to cells transfected with pRLnullΔ(77-1246)WTSPGE (negative control-HBV DNA coding for HBV Large, Middle, Small-HBsAg promoters, genes and enhancers I,II)9 in the corresponding real time PCR experiment (Fig. 4B). Thus, deletions in the spacer region of virus polymerase of mutant 313.1-1.86 mer and 761.1-1.86mer replicons do not affect virus replication.
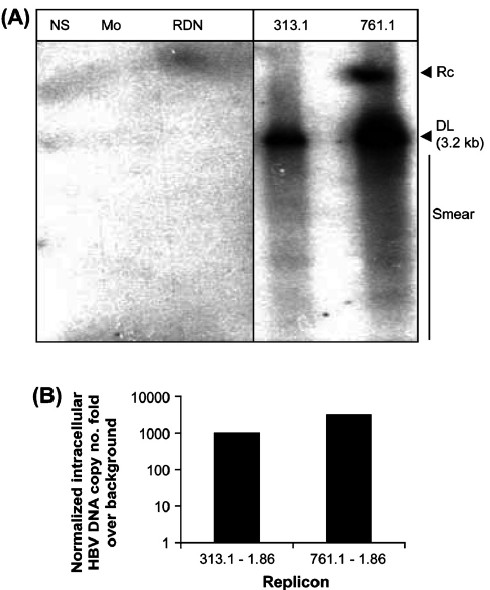
- (A) Huh7 cells transfected with the 313.1-1.86 and 761.1-1.86 constructs depicts Southern hybridization of intracellular DNA (Fig. 4A) at 72 h post-transfection. NS: Non specific DNA, Mo:Mock transfection, RDN: cells transfected with replication deficient negative control plasmid pRLnullΔ(77-1246)WTSPGE, M: 100pg marker consisting of linear double stranded fragments of HBV DNA, not shown, 313.1 : cells transfected with 313.1-1.86 construct, 761.1: cells transfected with 761.1-1.86 construct, RC: relaxed circular DNA. DL: double stranded linear DNA. The smear indicates HBV replication intermediates. (B) Relative levels of intracellular HBV DNA in Huh7 cells transfected with 313.1-1.86 and 761.1-1.86 constructs, by Taqman based quantitative real time PCR at 72 h post-transfection. The HBV DNA copy number fold over background normalized for transfection efficiency is depicted.
HBsAg secretion in co-transfection experiments of the wild-type genotype A and mutant genotype D replicons: During co-transfection of equimolar quantities of WT-1.86mer with mutant 313.1-1.86mer, 761.1-1.86mer replicons, the secretion of HBsAg was similar to the wild-type (Fig. 5).
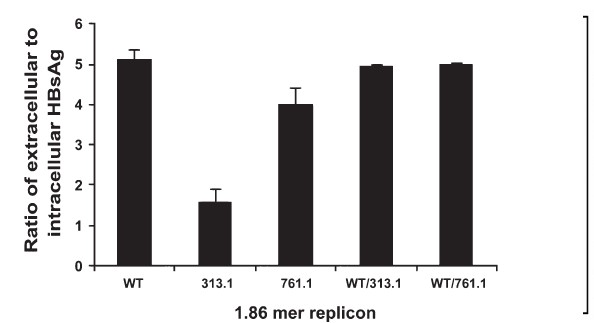
- Co-transfection of wild type and mutants. The indicated plasmids were transfected or co-transfected into Huh7 cells and the ratio of extracellular to intracellular HBsAg was calculated at 72 h post-transfection. Values are mean ± SEM for 3 observations.
Discussion
Hepatitis B virus isolates from two occult infection patients 2701 and 821 (AY161150-56, AY61161-63) were of genotype D without additional pre-S2/S promoter deletions5. HBV isolates (AY161147, AY161159) derived from occult infection patients 313 and 761 that showed reduced HBsAg secretion were of genotype D carrying additional pre-S2/S promoter deletions59. Since the contribution of the characteristic 33 nucleotide deletion of genotype D towards occult HBV infection is not known, we chose to analyse the role of this deletion in reducing HBsAg secretion by analyzing HBV isolates from occult infection patients 313(S5),761(S8) and 821(S9)59. In addition, we investigated the pre-S2/S promoter deletions described by us earlier5 for HBV isolates 313.1, 761.1 for various expression and replication parameters which can contribute to the occult phenotype.
Studies carried out earlier utilized expression constructs which consisted of the large surface (pre-S1) and small surface (pre-S2/S) promoters fused to reporters or the small surface gene59. Transient transfection of human hepatocellular carcinoma cell lines with these expression constructs may not reflect the exact biology seen during natural infection by HBV, which carries the full length genome.
Thus in the current study, we constructed HBV replicons, i.e. overlength 1.86mer/1.3mer constructs of wild-type genotype A and occult infection isolates of genotype D without and with pre-S2/S promoter deletions (821.1, 313.1,761.1).
Characteristic 33nt deletion of genotype D reduces HBsAg secretion: Comparing extracellular HBsAg between 1.3mer/1.86mer wild-type genotype A replicons and 821.1-1.3mer/1.86mer replicons of genotype D revealed that the 821.1-1.3mer/1.86mer replicons showed reduced extracellular HBsAg compared to wild-type genotype A. This suggests that the 33 nucleotide deletion reduces HBsAg export from the hepatocyte. The characteristic 33 nucleotide region of genotype A could be a possible target for a putative host microRNA, which needs experimental confirmation. MicroRNAs (miRNAs) are endogenous ~23 nt RNAs that pair with the mRNAs of protein-coding genes to direct their post-transcriptional repression2122. This mechanism of gene regulation occurs in animals and plants as well as animal viruses2123. It has been shown that the human microRNA hsa-miR-125a-5p interferes with expression of HBsAg24. We hypothesized that the 33 nt region of HBV pre-S1 mRNA is a possible target for a putative host microRNA. The host microRNA would function at the post-transcriptional level, by cleaving target mRNA or inhibit translation of mRNA. Absence of these 33 nucleotides in HBV genotype D pre-S1 mRNA would lead to higher levels of L-HBsAg. This would result in increased large HBsAg versus small HBsAg (L/S) ratio, intracellular accumulation and reduced secretion of HBsAg. Inputting the 33nt region of wild-type genotype A (X02763) as the target in the microRNA database, miRBase (www.mirbase.org)25 revealed a human microRNA hsa-mir-193b* (Accession: MIMAT0004767) (also known as hsa-mir-193-5p). Nucleotides 1 to 15 of this microRNA from the 5’ end could base pair with nt 2865 to nt 2879 in the pre-S1 mRNA region with a maximum free energy of -23.3 kcal/mole (RNA hybrid software version 2.2,26. The pre-miRNA of hsa-miR-193b* is present in Huh7 cells (source: Ferrolab, Datamining and bioinformatics group, 2007, miR-Ontology database, http://ferrolab.dmi.unict.it/miro/, search parameter: hsa-miR-193b* pre-miRNA in tissues). It has been shown that a genotype D isolate from surface antigen negative patient overexpressed pre-S1 mRNA and showed a concomitant secretion defect27.
Reduced HBsAg secretion in mutant 1.3mer genotype D replicons: Further reduction in extracellular HBsAg for the 313.1-1.3mer replicon was observed in comparison to 821.1-1.3mer, but this reduction was not significant. Deletions in the pre-S2/S promoter (Δnt 3026-3208) occur in 313.1-1.3mer which encompass positive regulatory regions B (nt 3077-3088), C (nt 3094-3127), D (nt 3130-3147), E (nt 3153-3172), G(nt 3177-3209) for S-HBsAg expression and the negative regulatory region F (nt 3065-3186) for S-HBsAg expression28. Also the ‘Z’ region (nt 3160-3221) which is a negative regulator of L-HBsAg expression29 is absent in 313.1-1.3mer leading to highly overexpressed L-HBsAg. Insufficient S-HBsAg is expressed from the mutated pre-S2/S promoters in the 313.1-1.3mer construct, which along with overexpressed L-HBsAg, should lead to highly increased L/S ratios. This should result in further intracellular accumulation and reduced extracellular HBsAg in the 313.1-1.3mer replicon in comparison to the 821.1-1.3mer replicon. Since the reduction in extracellular HBsAg in the 313.1-1.3mer replicon was insignificant in comparison to 821.1-1.3mer, we hypothesized that presence of two sets of enhancers in the 313.1-1.3mer replicon increased intracellular S-HBsAg levels which decreased L/S ratio leading to HBsAg secretion and extracellular HBsAg levels in the 313.1-1.3mer replicon similar to the 821.1-1.3mer replicon. We observed lesser extracellular HBsAg for the 761.1-1.3mer compared to the 313.1-1.3mer. Deletions of aa 63-81 in the overexpressed mutated L-HBsAg of 761.1-1.3mer could lead to misfolded L-HBsAg compared to properly folded L-HBsAg of 313.1-1.3mer where all the carboxy terminus amino acids 58-119 are deleted. Misfolded L-HBsAg of 761.1-1.3mer could aggregate with properly folded S-HBsAg leading to HBsAg aggregation and accumulation in cells.
Effect of two sets of pre-S1, pre-S2/S promoters and enhancers I,II on HBsAg secretion in case of the 1.86mer constructs: Using constructs consisting of a single set of surface antigen promoters, genes and enhancers it was previously shown that the L/S ratios for the wild-type genotype A was 0.179. This value increased to 1.89 (over ten-fold) for 313.1 and 0.56 (over three-fold) for 761.1, resulting in reduced HBsAg secretion for these constructs relative to wild-type genotype A. In the present study, extracellular HBsAg was similar in the 821.1-1.86mer and 313.1-1.86mer replicons. In the 313.1-1.86mer L-HBsAg is overexpressed due to presence of the 33nt deletion in the pre-S1 ORF1, and absence of the ‘Z’ region in the pre-S2/S promoter. Insufficient S-HBsAg is expressed from the mutated pre-S2/S promoters in the 313.1-1.86mer construct, which along with overexpressed L-HBsAg, leads to highly increased L/S ratios. However, along with an increase in L-HBsAg, presence of two sets of enhancers and two sets of pre-S2/S promoters in the 313.1-86mer construct could also elevate the levels of S-HBsAg. This would result in L/S ratios greater than that of genotype A but similar to that of 821.1-1.86mer, leading to extracellular HBsAg values for 313.1-1.86mer replicon lesser than that of wild-type genotype A but similar to 821.1-1.86mer. Replicon 761.1-1.86mer showed reduction in HBsAg secretion compared to the wild-type genotype A, 1.86mer replicon. Deletions in the pre-S2/S promoter (Δnt 3039-3095), encompassing only the positive regulatory region B for S-HBsAg expression2829 are present. This results in reduction of S-HBsAg, increased L/S ratio, intracellular HBsAg accumulation, and reduced extracellular HBsAg. Extracellular HBsAg in the 761.1-1.86mer replicon was greater than the 313.1-1.86mer replicon. Positive regulatory region B for S-HBsAg expression was deleted in the 761.1-1.86mer replicon as compared to deletion of positive regulatory regions B,C,D,E,G and negative regulatory region F for S-HBsAg expression in case of the 313.1-1.86mer replicon2829. Region Z is absent in the 313.1-1.86mer replicon but present in the 761.1-1.86mer replicon. Together these deletions lead to greater L/S ratio of 313.1-1.86mer compared to 761.1-1.86mer resulting in lesser extracellular HBsAg for 313.1-1.86mer.
Replication of 313.1-1.86 and 761.1-1.86 replicons: In case of the 313.1-1.86mer replicon which carries deletions in amino acids 185 to 195, 240 to 300 of the virus polymerase, the presence of intracellular HBV DNA by Southern blot analysis and real time PCR was observed. The quantity of intracellular virus DNA in the 313.1-1.86mer replicon is thousand fold greater than cells transfected with pRLnullΔ(77-1246)WTSPGE negative control. It is concluded that changes in the polymerase in mutant 313.1 do not affect HBV replication. These results agree with a report where it was found that the HBV polymerase function was not affected in Huh7 cells by mutants with deletions from nt 2914 to nt 2957, and nt 2989 to nt 3070 (which correspond to the pre-S2/S promoter region) or both30. The same group showed that an in frame deletion of 183 nucleotides in the pre-S1 region (nucleotide 2983 and 3167 i.e. amino acid 47 to 107 of the large envelope protein) would generate a deleted large envelope protein that is retained in the endoplasmic reticulum, leading to an accumulation of nucleo-capsids containing virus DNA31.
Changes in the overlapping polymerase open reading frame of the 761.1-1.86mer replicon, which carries deletions of amino acids 185 to 195, 243 to 261 did not affect virus replication as indicated by presence of virus DNA by Southern blot analysis and virus DNA copy number values obtained in real time PCR.
Co-transfection experiments: Co-transfection experiments of equimolar quantities of the wild-type 1.86mer replicon with mutant replicons, 313.1-1.86 and 761.1-1.86 resulted in HBsAg secretion similar to the wild-type. This indicated that S-HBsAg was produced by the wild-type in excess, resulting in normal L/S ratios that could overcome the secretion defect in case of the mutants due to abnormal L/S ratios.
In conclusion, it has been demonstrated that the characteristic 33nt deletion of genotype D, HBV contributes to reducing HBsAg secretion in occult infection patients, infected with this genotype. This conclusion is validated by observations that amplification of the HBV pre-S1 gene from 57 occult infection patients revealed that all the patients carried the characteristic 33 nucleotide deletion of genotype D13. Secondly it has also been shown that, least quantities of secreted HBsAg were observed in HBV genotype D10. Additional pre-S2/S promoter deletions may worsen this reduced HBsAg secretion phenotype. This observation is validated by the presence of naturally occurring deletions in the pre-S2/S promoter region in several cases of occult HBV infection56. Concomitant changes in the overlapping polymerase open reading frame do not affect virus replication. Occult hepatitis B infection is a problem in many parts of the world including India, which affects blood banking and diagnosis of chronic hepatitis B. Faulty blood banking practices and diagnosis of chronic hepatitis B may result in transfusion transmitted hepatitis and also in hepatitis during solid organ transplantation32. Occult hepatitis B is also implicated in hepatocellular carcinoma33. Hence, the present study underscores the need to check for the presence of serum HBV DNA, especially in cases of HBsAg negative blood34, prior to blood transfusion. Additionally, HBV needs to be genotyped in occult infections. The present study demonstrates the biology of HBV in occult infection patients infected with genotype D HBV. This may help to develop rational therapies in case of such infections, for example, targeted drug delivery to the liver where the virus resides.
References
- Statements from the Taormina expert meeting on occult hepatitis B virus infection. J Hepatol. 2008;49:652-7.
- [Google Scholar]
- Occult hepatitis B virus infection: a covert operation. J Viral Hepat. 2010;17:1-15.
- [Google Scholar]
- Occult hepatitis B virus infection in chronic liver disease: full length genome and analysis of mutant surface promoter. Gastroenterology. 2004;127:1356-71.
- [Google Scholar]
- Occult hepatitis B virus infection in hepatitis B vaccinated children in Taiwan. J Hepatol. 2009;50:264-72.
- [Google Scholar]
- Prevalence and significance of hepatitis B virus (HBV) pre-S mutants in serum and liver at different replicative stages of chronic HBV infection. Hepatology. 2001;33:277-86.
- [Google Scholar]
- High prevalence and mapping of pre-S deletion in hepatitis B virus carriers with progressive liver diseases. Gastroenterology. 2006;130:1153-68.
- [Google Scholar]
- Role of surface promoter mutations in hepatitis B surface antigen production and secretion in occult hepatitis B virus Infection. J Med Virol. 2007;79:220-8.
- [Google Scholar]
- Influence of hepatitis B virus genotypes on the intra- and extracellular expression of virus DNA and antigens. Hepatology. 2006;44:915-24.
- [Google Scholar]
- Prevalence and virological features of occult hepatitis B virus infection in female sex workers who work uncontrolled in Turkey. Liver Int. 2009;29:227-30.
- [Google Scholar]
- Occult hepatitis B virus infection in a cohort of HIV-positive patients: correlation with hepatitis C virus coinfection, virological and immunological features. Infection. 2009;37:445-9.
- [Google Scholar]
- Hepatitis B virus genotype, HBsAg mutations and co-infection with HCV in occult HBV infection. Clin Res Hepatol Gastroenterol. 2011;35:554-9.
- [Google Scholar]
- The role of envelope proteins in hepatitis B virus assembly. Proc Natl Acad Sci USA. 1991;88:1059-63.
- [Google Scholar]
- A preS mutation isolated from a patient with chronic hepatitis B infection leads to virus retention and misassembly. Gastroenterology. 1997;113:1976-82.
- [Google Scholar]
- The genome of hepatitis B virus contains a second enhancer: cooperation of two elements within this enhancer is required for its function. J Virol. 1990;64:4281-7.
- [Google Scholar]
- Dysregulated surface gene expression from disrupted hepatitis B virus genomes. J Virol. 1993;67:7032-40.
- [Google Scholar]
- Enhancer I predominance in hepatitis B virus gene expression. Mol Cell Biol. 2004;24:1799-808.
- [Google Scholar]
- Quantitation of hepatitis B virus genomic DNA by real-time detection PCR. J Clin Microbiol. 1999;37:2899-903.
- [Google Scholar]
- Analysis of genomic DNA by southern hybridization. In: Molecular cloning a laboratory manual Vol II. (2nd ed). New York: Cold Spring Harbor Laboratory Press; 1989. p. :9.31-9.58.
- [Google Scholar]
- Human microRNA hsa-miR-125a-5p interferes with expression of hepatitis B virus surface antigen. Nucleic Acids Res. 2011;39:5157-63.
- [Google Scholar]
- Characterization of two hepatitis B virus populations isolated from a hepatitis B surface antigen-negative patient. Hepatology. 2002;35:1215-24.
- [Google Scholar]
- Complex regulation of transcription from the hepatitis B virus major surface antigen promoter in human hepatoma cell lines. J Virol. 1991;65:4805-11.
- [Google Scholar]
- Negative regulation of the hepatitis B virus pre-S1 promoter by internal DNA sequences. Virology. 1989;170:251-60.
- [Google Scholar]
- Properties of hepatitis B virus pre-S1 deletion mutants. Virology. 1994;199:292-300.
- [Google Scholar]
- The small envelope protein is required for secretion of a naturally occurring hepatitis B virus mutant with pre-S1 deleted. J Virol. 1997;71:5449-54.
- [Google Scholar]
- Hepatitis B virus infection and transfusion medicine: science and the occult. Transfusion. 2008;48:1001-26.
- [Google Scholar]
- Association between occult hepatitis B infection and the risk of hepatocellular carcinoma: a meta-analysis. Liver Int. 2012;32:231-40.
- [Google Scholar]
- Nucleic acid testing to detect HBV infection in blood donors. N Engl J Med. 2011;364:236-47.
- [Google Scholar]




