
Translate this page into:
MicroRNA expression signature in gastrointestinal stromal tumour & their molecular & histological features
For correspondence: Dr Kim Vaiphei, Department of Histopathology, Postgraduate Institute of Medical Education and Research, Chandigarh 160 012, India e-mail: kvaiphei2009@gmail.com
-
Received: ,
Abstract
Background & objectives
In gastrointestinal stromal tumour (GIST), not only genetic abnormalities are responsible for adverse clinical events, but epigenetic modifications also play a crucial role. MicroRNA (miRNA) dysregulation plays a significant role in carcinogenesis as miRNAs serve as natural silencer for their targets. Our study aimed to explore the miRNAs expression and its association with molecular and histopathological characteristics of GIST.
Methods
Fifty GIST samples, including 45 formalin fixed paraffin embedded (FFPE) and fresh tissues were included. Peripheral non-tumour tissues were used as controls. All the cases were confirmed using immunohistochemistry. RNA was extracted using miRNA-specific kit, and the expression was performed using RT-qPCR. The data were evaluated using AriaMx software version 1.5 (Agilent, US). MiRNAs expression was analyzed by using the relative quantification method (ΔΔCT).
Results
miR-221, miR-222, miR-494 and miR-34a showed significant down-regulation in tumours relative to non-tumour tissues. The expression levels of these miRNAs were significantly down-regulated in c-KIT (proto-oncogene encoding the tyrosine kinase transmembrane receptor)-positive tumours compared to c-KIT-negative. Further analysis revealed that reduced expression was associated with spindle subtypes and gastric localization. However, there was no significant correlation with other histological features. Additionally, miR-221/222, and miR-494 were down-regulated in most of the KIT exon 11 mutant subtypes, while miRNA-34a was associated with platelet derived growth factor receptor alpha (PDGFRA) mutations.
Interpretation & conclusions
The present study showed that the down-regulation of these miRNAs may help better molecular classification and characterization of GISTs. Our results offer new insight into the association between miRNAs and histological features, enabling a more thorough understanding of GISTs at the molecular level.
Keywords
Gastrointestinal stromal tumour
microRNAs
epigenetic
real time-PCR
histopathology

Gastrointestinal stromal tumours (GISTs) are considered as heterogeneous entities, commonly characterized by activating proto-oncogene encoding the tyrosine kinase transmembrane receptor (KIT) and platelet derived growth factor receptor alpha (PDGFRA mutations1. GISTs are determined to express KIT and CD34, which are the distinguishing characteristics of Interstitial Cells of Cajal (ICC) since it is believed that ICC are the source of GISTs2. The diagnoses of GISTs have become more precise since the discovery of CD117 and DOG1. GISTs are most typically found in the stomach and small intestine, with just around five per cent found in the colon or rectum and about two per cent detected in the oesophagus or other organs3.
MicroRNAs (miRNAs) are recently identified non-protein-coding RNA molecules consisting of 19 to 24 nucleotides4. Post-transcriptionally, miRNAs control gene expression by restricting protein translation or by degrading target molecules5. They serve as natural silencers for the target transcripts and are involved in virtually all cellular processes, including cell division, differentiation, and even apoptosis. Notably, miRNA expression has also been identified as vulnerable to the regulation of the epigenetic process, and miRNA-based therapeutics are currently under investigation.
The involvement of functional connections among miRNAs and KIT in GISTs has been shown in numerous studies. It has been found that miR-221 and miR-222 expressions coincide inversely with KIT expression, indicating that these microRNAs may negatively impact the KIT expression6. It has also been reported that the components of miR-222/221 clusters and miR-494 target KIT and ETV17,8. This implies that these miRNAs may have therapeutic value in managing GISTs. Increased miR-221 and miR-494 production in KIT-positive cells inhibit cell proliferation by downregulating KIT signalling transduction pathways9. Further, these were discovered to be downregulated in both WT-type and mutant GISTs7. The expression of miR-221/222 was discovered to be considerably lower in GIST when compared to other similar tumours and control tissues10.
Kim WK and colleagues8 discovered that miR-494 was a negative regulator of KIT in GIST. Furthermore, the researchers found that miR-494 over-expression in GISTs may be a viable method of treatment. According to the findings of the other research team, miR-34a decreased the expression of several target genes, particularly PDGFRA10,11. RNA interference-induced inhibition of PDGFRA in GIST-T1 cells decreased cell proliferation, indicating that the anticancer effect of miR-34a was partially mediated by PDGFRA targeting10,11.
However, a sufficient amount of pertinent data has not yet been collected. As a result, it is probable that, in addition to conventional therapy, a variety of medicines with unique modes of action and targeting distinct molecular mechanisms will be investigated and potentially added to the GIST treatment arsenal12. Various studies show the potential contribution of epigenetic variables to the pathophysiology and prognosis of GIST. It is possible that the evolution of GIST is the product of both epigenetic and genetic events13,14. We need to learn more about these molecular pathways because it could help us make better treatments.
There is accumulating evidence that complex epigenetic modifications occur throughout the development of GIST and correspond with an increase in tumour aggressiveness. Understanding these genetic alterations may help predict clinical behaviour and provide information about responsiveness to or resistance to therapeutic strategy. In addition to serving as possible therapeutic agents or targets, regulators of epigenetic pathways might also act as therapeutic agents or targets. Before drawing definitive conclusions, additional studies must be conducted to shed light on these aspects.
Material & Methods
The study was conducted at the department of Histopathology, Post-graduate Institute of Medical Education and Research (PGIMER), Chandigarh, India from January 2012 to December 2020 after obtaining the ethical clearance from the Institutional Ethics Committee.
Sample processing
Formalin-fixed and paraffin-embedded (FFPE) blocks of the initial 58 cases were obtained from the department of Histopathology, PGIMER; however, complete repeated immunocytochemical analysis and miRNA profiling were possible in 45 cases; 13 cases could not pass the quality metrics for miRNA amplification, so they were excluded from the study.
Clinically and histologically diagnosed GIST affected individuals over eight-year period (2012 to 2020) were included for miRNA expression analyses. The cases were from department of surgery, PGIMER, who had presented with abdominal mass. Fifty GIST samples including 45 FFPE and five fresh tissues were included. Peripheral non-tumour tissues were used as controls. Among these, FFPE specimens also were previously used for molecular analysis in KIT (exons 9, 11, 13 and 17) and PDGFRA (exons 12, 14 and 18) for diagnosis. However, the mutational data were available in 42 GIST cases (Supplementary Table). The pathological information of each case was reviewed including cell morphology, primary tumour site, tumour size, mitotic index and risk grade.
Diagnosis and screening
After establishing GIST diagnosis from the surgically resected specimens, frozen samples were subjected to study the composition of the tissue. All surgically resected specimens were processed for routine diagnosis purposes; hematoxylin and eosin staining and immunohistochemistry. Light microscopic review and immunohistochemical analysis for CD117 [Clone, Rabbit polyclonal anti-CD117 antibodies (CD 117, Dako, Denmark) at dilution 1:200] were performed and reviewed independently without knowledge of previous clinical information. One of the limitations of this study was that we did not test PDGFRA and DOG-1 immunostaining in any GIST case. We based our diagnosis on CD117 staining.
RNA extraction from FFPE and fresh tissues
Total RNA was isolated from FFPE and fresh tissue using miRNA-specific kit (Qiagen, Germany) Adjacent control tissues, displaying no macroscopic abnormalities, were meticulously chosen for comparison with the tumour samples. Surgical margins, ranging from 5 to 7 cm away from the primary tumour sites, were excised and meticulously evaluated through histopathological examination to ensure the absence of bias in the control samples. The frozen samples were ground to fine particles with the help of a metal spatula, and the powdered tissue was scraped and mixed with Lysis Buffer. The FFPE blocks having sufficient tumours were selected for RNA isolation. Using a standard microtome, 2-3 sections of FFPE tissue with a thickness of 5 µm were cut and immediately deparaffinized using 300 μl of deparaffinization solution (Qiagen, Germany). The tissue was digested in 150 μl of PKD buffer and10μl Proteinase K (Qiagen) and incubated at 56°C for overnight digestion followed by 80°C incubation for 15 min. Manufacturer’s protocol was followed for miRNA extraction. The RNA quantity and quality were checked by measuring absorbance in a nanodrop (NanoDrop 2000, ThermoScientific, USA) at 260 nm (1 O.D. = 40 μg/ml); A260/A280 ratio of 2 was considered purified RNA and A260/A230 ratio >1.7 considered as highly purified RNA.
cDNA synthesis
Following the protocol outlined by the manufacturer of the miScript II RT cDNA synthesis kit (Qiagen, Germany), 100 ng of RNA was subjected to reverse transcription in a total volume of 20 μl. The concentrated RNA was diluted in RNase-free water to make the final concentration of 1μg of RNA used for cDNA synthesis. Master mix for reverse transcription was prepared on ice in accordance with the stated instructions. The final incubation at 37°C for 60 min was followed by the termination of the reaction by incubating for 5 min at 95°C. The products of the reverse transcription reaction were either utilized immediately for qPCR or kept at -20°C for further use.
miRNA amplification using real-time qPCR
The expression levels of microRNAs were assayed by Quantitative RT-PCR analysis performed on AriaMX Real-time PCR system (Agilent, US). About 50 ng of cDNA was used in each sample with the SYBRGreen (Agilent, US). The qPCR master mix was prepared in a final volume of 10 μl using 5 μl of SYBRGreen, 0.5 μl of forward and reverse primer, 50 ng/μl of cDNA and nuclease-free water in 96 well plates on ice. The plate was covered with sealing film and centrifuged at 1000 g/1 min at 4°C to eliminate the bubble that could interfere with PCR reactions. All samples were evaluated in triplicates using the following cycling conditions: 40 cycles of initial denaturation for 10 sec at 95°C, 15 sec of annealing at 58°C, and extension at 72°C for 20 seconds were performed after the initial denaturation for 5 min at 95°C. The melting curve conditions were 95°C for 10 sec, 5°C (+annealing temp) for 1 min and 95°C for 20 sec. The details of miRNA primer sequences are depicted in Supplementary Table.
Analysis of real-time qPCR data
Following the generation of a melting curve for each test in each run to determine the precise amplification efficiency, miRNAs expressions in non-tumour and tumour tissue samples were statistically evaluated and adjusted against RNU6 levels for all samples. Positive PCR control and negative control were also used. AriaMx software version 1.5 (Agilent, US) was used to evaluate the data obtained as a threshold cycle (CT value). The baseline was automatically assigned for all wells in the software to calculate threshold values above background noise. A negative call was deemed to be a CT value of less than 35. Each miRNAs expression profiling data was analysedusing a relative quantification method, delta-delta Ct method (ΔΔCT)13. The miRNA of interest (MOI) and the U6 RNA were amplified in each sample to determine the Ct value for each.
Statistical Analysis
miRNAs expression levels were calculated and compared with clinicopathological characteristics i.e., immunostaining, tumour site, and cell morphology as a factor using the Kruskal-Wallis rank sum test and Wilcoxon rank sum exact test. The P≤0.05 was considered as a significant difference and the pair-wise Wilcoxon rank sum exact test was applied if the Kruskal-Wallis rank sum test was found significant. The box plot was chosen to create a distributional (i.e., variance, interquartile range and outliers) overview for each miRNA expression level. A post-hoc test of Tukey’s was applied as a correction to calculate the final P value. The analysis was performed on the R Studio base package and plotted using the R package “ggpubr”.
Results
Sample characteristics
Deregulation of miRNA expression is considered to be one of the most significant factors contributing to carcinogenesis. We tried to focus on the previously described four miRNAs including miR-222 and miR-494, miR-221 and miR-34a in 50 GISTs specimens. Four potential miRNAs were selected because of their statistically significant correlation with KIT expression. MiR-222 and miR-221, miR-494 have been suggested to be targeting the KIT gene while miR-34a targets PDGFRA. We investigated the miRNAs’ expression in GIST and contrasted the variations with adjacent peripheral non-tumour tissue, clinicopathological characteristics and molecular alterations.
The study participant’s clinicopathological characteristics are depicted in Table I. All the study participants were diagnosed based on c-KIT immunostaining. Among these, 45 cases showed positivity for CD117, whereas five cases were negative. Of the 50 cases, eight study participants developed metastasis, most commonly in liver (n=6) and one case each in lymph node and omentum, respectively. Seven study participants presented with recurrent tumours. In c-KIT negative cases, one patient with jejunal involvement exhibited regional lymph node metastasis.
|
Sample code |
Sex/age (yr) |
CD117 | Tumour site |
Tumour size (cm) |
Mitotic index (/50HPF) |
Cell morphology | Tumour risk |
Mutational status |
Metastasis |
|---|---|---|---|---|---|---|---|---|---|
| 00G1 | M/26 | Positive | Stomach | <11 | 5/50HPF | Spindle | High | Wild type | Yes |
| 00G2 | F/55 | Positive | Stomach | <13 | 3/50HPF | Spindle | High | K-11 | No |
| 00G3 | F/68 | Positive | Stomach | <8 | NM | Spindle | NM | K-11 | No |
| 00G4 | M/45 | Positive | S. intestine | <5.5 | 2/50HPF | Spindle | Low | K-9 | No |
| 00G5 | F/57 | Negative | S. intestine | <12 | NM | Spindle | High | K-13 | No |
| 00G6 | F/50 | Negative | Rectum | <13 | 10/50HPF | Spindle | High | P-12 | Yes |
| 00G7 | M/57 | Negative | Stomach | <6.5 | NM | Epithelioid | Intmd. | P-18 | No |
| 00G8 | M/65 | Positive | Stomach | <6 | 3/50HPF | Spindle | Intmd. | No | Yes |
| 00G9 | M/50 | Positive | Stomach | <5 | 5/10HPF | Spindle | Intmd. | K-11 | Yes |
| 00G10 | F/38 | Positive | S. intestine | <10 | 2/50HPF | Spindle | Intmd. | K-9 | No |
| 00G11 | M/54 | Positive | Omentum | <19 | NM | Epithelioid | High | K-11 | No |
| 00G12 | M/67 | Positive | Stomach | <7 | 3/50HPF | Spindle | Intmd. | K-11 | No |
| 00G13 | F/46 | Positive | Stomach | <16 | NM | Epithelioid | High | K-11 | Yes |
| 00G14 | M/65 | Positive | Colon | NM | 1/50HPF | Spindle | Low | K-11 | Yes |
| 00G15 | F/50 | Positive | Stomach | NM | NM | Mixed | NM | K-9 | No |
| 00G16 | M/34 | Positive | Stomach | NM | NM | Spindle | NM | P-18 | No |
| 00G17 | F/37 | Positive | S. intestine | <13 | 10/50HPF | Mixed | High | K-17 | Yes |
| 00G18 | F/34 | Positive | S. intestine | NM | NM | Spindle | NM | K-11 | No |
| 00G19 | M/50 | Positive | Mesentery | <11 | 48/10HPF | Epithelioid | High | K-11 | Yes |
| 00G20 | M/53 | Positive | Stomach | <0.4 | NM | Spindle | Low | K-11 | No |
| 00G21 | M/61 | Positive | Stomach | <4 | 2/50HPF | Spindle | Low | K-11 | No |
| 00G22 | F/54 | Positive | Stomach | NM | NM | Spindle | NM | K-11 | No |
| 00G23 | M/60 | Positive | S. intestine | NM | 1/50HPF | Spindle | Low | No | No |
| 00G24 | F/67 | Positive | Stomach | <4 | 10/50HPF | Spindle | High | K-11 | No |
| 00G25 | F/48 | Positive | Stomach | <12 | <2/10HPF | Epithelioid | High | K-11 | No |
| 00G26 | M/50 | Positive | S. intestine | <8 | <5/20HPF | Spindle | Intmd. | K-11 | Yes |
| 00G27 | F/58 | Negative | Stomach | <7 | NM | Spindle | NM | K-9 | No |
| 00G28 | M/57 | Positive | S. intestine | NM | NM | Spindle | NM | K-11 | No |
| 00G29 | F/45 | Positive | Stomach | <12 | NM | Spindle | High | K-17 | Yes |
| 00G30 | F/34 | Positive | Stomach | NM | NM | Spindle | NM | K-11 | No |
| 00G31 | M/27 | Positive | Stomach | NM | NM | Epithelioid | NM | K-11 | No |
| 00G32 | M/56 | Positive | Stomach | NM | NM | Spindle | NM | K-11 | No |
| 00G33 | M/32 | Positive | S. intestine | NM | NM | Spindle | NM | K-17 | No |
| 00G34 | F/48 | Positive | S. intestine | <12 | <2/10HPF | Spindle | High | K-11 | No |
| 00G35 | F/49 | Positive | S. intestine | NM | NM | Spindle | NM | P-18 | No |
| 00G36 | M/21 | Negative | Stomach | <17 | 1/50HPF | Spindle | High | P-18 | Yes |
| 00G37 | M/50 | Positive | S. intestine | NM | NM | Spindle | NM | No | No |
| 00G38 | F/28 | Positive | S. intestine | <7 | 3/50HPF | Spindle | Low | K-17 | Yes |
| 00G39 | F/67 | Positive | S. intestine | NM | NM | Spindle | NM | K-11 | No |
| 00G40 | F/75 | Positive | Stomach | <8 | 8/50HPF | Spindle | Intmd. | K-11 | Yes |
| 00G41 | M/43 | Positive | Colon | <13 | NM | Spindle | High | No | Yes |
| 00G42 | M/71 | Positive | Stomach | NM | NM | Epithelioid | NM | K-11 | No |
| 00G43 | M/36 | Positive | Stomach | <12 | >5/50HPF | Mixed | High | K-11 | Yes |
| 00G44 | M/48 | Positive | Stomach | <15 | NM | Epithelioid | High | K-11 | No |
| 00G45 | M/67 | Positive | Stomach | NM | NM | Spindle | NM | K-9 | No |
| 00G46 | M/57 | Positive | Stomach | NM | NM | Spindle | NM | No | No |
| 00G47 | M/45 | Positive | Stomach | <7 | <5/50hpf | Spindle | Intmd. | No | No |
| 00G48 | M/38 | Positive | S. intestine | NM | NM | Spindle | NM | No | No |
| 00G49 | F/48 | Positive | Stomach | NM | NM | Spindle | NM | No | No |
| 00G50 | M/50 | Positive | Stomach | <6 | 8/10hpf | Spindle | Intmd. | No | No |
S. Intestine, small intestine; NM, data not mentioned; No, no metastasis and mutations; Intmd., intermediate risk; F, female; M, male
Based on the morphological analyses, 39 cases showed spindle cell morphology followed by epithelioid (n=8) and the remaining three were mixed type. The overall tumour size ranged from 3 to 19 cm. The large tumour size (>10 cm) were found in the majority of the cases (n=16), while intermediate tumour size (>5-<10 cm) was reported in 12 cases. However, small size tumours (<5 cm, n=5) were fewer. The presentation of GISTs varies depending on the primary tumour location with differing prognostic and predictive relevance. Most primary GISTs had gastric localization (n=30) followed by the small bowel (n=15) and the remaining were found in the Colon (n=2), rectum (n=1), omentum (n=1) and mesentery (n=1).
Mitotic count is among the most crucial aspects of pathological examination for determining the likelihood of tumour risk. The mitotic rate ranged between 2-5/50HPF and was reported in the majority of the cases (n=16) followed by 5-10/50HPF in six cases, and the higher mitotic rate (>10/50HPF) was found only in one case. The patients were classified according to the NIH standard criteria for determining the probability of aggressive clinical progression of primary GISTs. The findings of our study revealed that a total of 16 cases came under the high risk category, nine cases as intermediate, and six cases were categorized under the low-risk category. Molecular data were available in 41 cases in which 25 cases had KIT exon 11 mutations followed by exon 9 (n=5), Exon 13 (n=1) and 17 (n=4) while PDGFRA exon 12 and 18 mutations were reported in one and four cases, respectively.
MiRNA expression in GIST tumour and normal tissue
In our study, we performed real-time PCR (RT-qPCR) analyses to assess the expression levels of miR-221, miR-222, miR-494 and miR-34a between tumours and peripheral non-tumour tissue. We observed that in the majority of the cases, there was a significantly reduced expression in miR-221 (P=0.009), miR-222 (P=0.012), miR-494 (P=0.015), miR-34a (P=0.024), in tumour tissues relative to non-tumour (P=0.48) (Fig. 1).
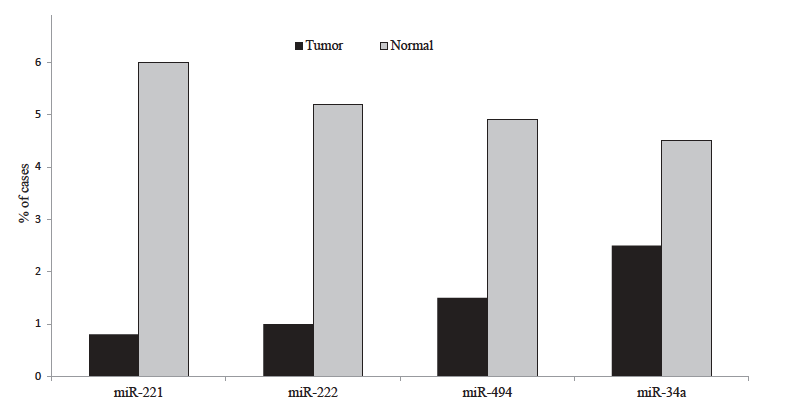
- miRNA expression level in tumour and non-tumour tissue. miR-221 and miR-222, miR-494 and miR-34a showing considerable difference in expression level between GIST tumour and pheripheral non-tumour tissue (median expression level, 0.015).
MicroRNAs expression and its correlation with histological features
miRNA expression levels in KIT-negative and KIT-positive tumours exhibited a substantial difference (Fig. 2A). We found a significant correlation (P=0.00000008) between c-KIT and miR-221, miR-494, miR-222, and miR-34a expression pattern, especially between KIT positive and negative tumours. The miR-221, miR-494, miR-34a and miR-222 expression levels, were significantly down-regulated in KIT-positive tumours whereas, KIT-negative tumours displayed the exact opposite expression pattern. However, there were only five KIT-negative cases in our study. Based on our study, this inverse relationship demonstrated that these miRNAs directly or indirectly regulated KIT expression and might be the modulators of KIT in GISTs. Further, we explored the clinical importance of miRNA expressions in histological subtypes of GIST. MiR-221, miR-494, miR-222 and miR-34a were found to be down-regulated in the majority of the spindle subtype; however, up-regulation was reported in a few cases. These miRNAs did not show a significant correlation with epithelioid and mixed sub-types; these cases were very few.
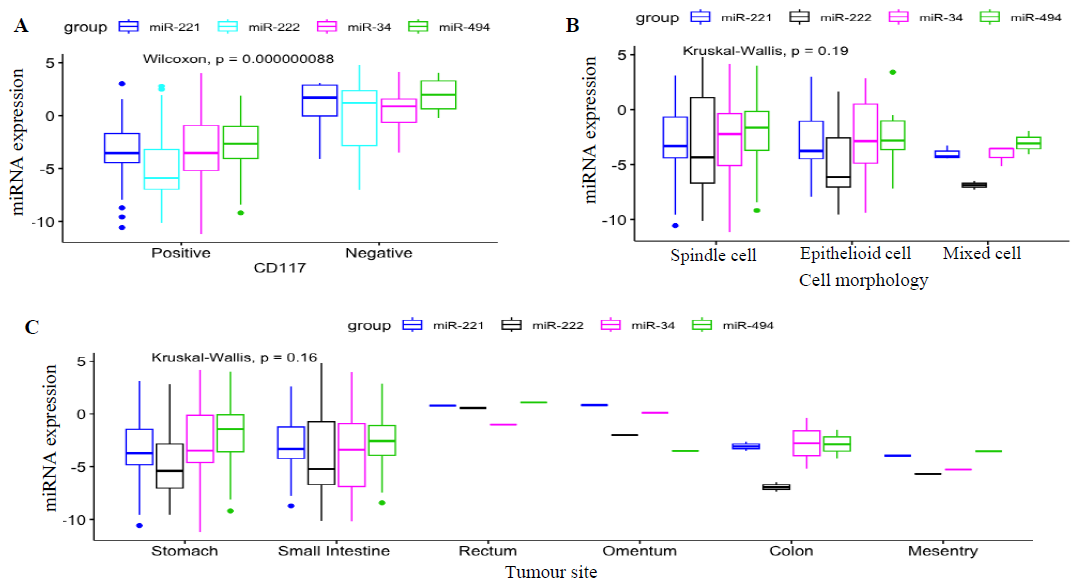
- miRNA expression level and its association with histological features. (A) KIT immunostaining: miRNAs expression levels in KIT-negative and KIT-positive tumours exhibited a substantial difference (P value: 0.00000008). miRNAs were significantly down-regulated in KIT-positive tumours whereas, KIT-negative tumours displayed the exact opposite expression pattern as KIT-positive tumours. (B) Cellular morphology: miRNAs were found down-regulated in the majority of the spindle subtype (P value=0.16). However, these did not showed any significant correlation with epithelioid and mixed sub-types. (C) Primary tumour sites: majority of the gastric and small intestinal GIST showed increased expression level (P value=0.19) however; there were no significant correlation with other primary tumour.
Moreover, all these miRNAs showed up-regulation in a few cases, irrespective of various histological parameters. Based on the overall statistical evaluation, the Kruskal-Wallis rank sum test did not find a significant association between miRNA and cellular morphology (P=0.19; Fig. 2B).
Further, we attempted to identify the correlation between miRNAs and the site of tumour occurrence. The miRNA-data were analyzedamong gastric, small intestinal, colon, rectum, omentum and mesentery. MiR-221, miR-494, miR-34a and miR-222 were found down-regulated in the majority of the gastric and small bowel GIST; however, there was no positive correlation in the colon, rectum, omentum and mesentery. Based on the Kruskal-Wallis rank sum test, overall expression showed a non-significant association with tumour sites (P=0.16; Fig. 2C).
MiRNA and its correlation with molecular subtypes
To categorize different subtypes of GISTs based on miRNA expression profiles, molecular analysis was carried out using relevant gene sets and exonal mutation with miRNA expression. The miRNA-data correlated with KIT exon 9, 11 and 17 and PDGFRA exon 18. We found a significant correlation between KIT exons 11 and 17 mutant subtypes and miRNA expression. Further analysis revealed that miR-221 and miR-222, miR-494 expression level were greatly reduced in KIT Exon 11 and 17 mutated GIST, most commonly reported in Gastric GIST(n=15) followed by small intestine (n=6).Similarly, miRNA-34a was found down-regulated in PDGFRA exon 18 mutant subtypes most frequently in small bowel GIST (n=3).However, there was no significant correlation found in KIT exon 9 and PDGFRA exon 18 and miR-222, miR-221, miR-494 expression in the small intestine and Gastric GIST. MiRNAs and its correlations with mutational subtypes and primary tumour sites are depicted in Table II.
| miRNAs | KIT mutation | PDGFRA | ||||||
|---|---|---|---|---|---|---|---|---|
|
Exon 9 (n=5) |
Exon 11 (n=25) |
Exon 17 (n=4) |
Exon 18 (n=4) |
|||||
|
Down- regulation |
Up- regulation |
Down- regulation |
Up- regulation |
Down- regulation |
Up- regulation |
Down- regulation |
Up- regulation |
|
| miR-221 | n=3 | n=2 | n=22 | n=3 | n=4 | --- | n=2 | n=2 |
| Tumour sites |
S.bowel (n=1) Stomach (n=2) |
S.bowel (n=1) Stomach (n=1) |
S.bowel (n=6) Stomach (n=15) Colon (n=1) |
S.bowel (n=1) Stomach (n=2) |
S.bowel (n=2) Stomach (n=2) |
--- |
S.bowel (n=1) Stomach (n=1) |
S.bowel (n=1) Stomach (n=1) |
| miR-222 | n=3 | n=2) | n=21 | n=4 | n=4 | --- | n=1 | n=3 |
| Tumour sites |
S.bowel (n=1) Stomach (n=2) |
S.bowel (n=1) Stomach (n=1 |
S.bowel (n=6) Stomach (n=15) |
S.bowel (n=1) Stomach (n=3) |
S.bowel (n=2) Stomach (n=2) |
--- | S.bowel (n=1) |
S.bowel (n=2) Stomach (n=1) |
| miR-494 | n=2 | n=3 | n=23 | n=2 | n=4 | --- | n=1 | n=3 |
| Tumour sites |
S.bowel (n=1) Stomach (n=1) |
S.bowel (n=1) Stomach (n=2) |
S.bowel (n=6) Stomach (n=15) Mesentry (n=1) |
S.bowel (n=1) Stomach (n=1) |
S.bowel (n=2) Stomach (n=2) |
--- | Stomach (n=1) |
S.bowel (n=2) Stomach (n=1) |
| miR-34a | n=3 | n=2 | n=21 | n=4 | n=3 | n=1 | n=4 | --- |
| Tumour sites |
S.bowel (n=1) Stomach (n=2) |
Stomach (n=2) |
S.bowel (n=5) Stomach (n=16) |
S.bowel (n=2) Stomach (n=2) |
S.bowel (n=2) Stomach (n=1) |
S.bowel (n=1) |
S.bowel (n=3) Stomach (n=1) |
--- |
S. bowel, small bowel
Discussion
GISTs are considered heterogeneous entity, and not only genetic abnormalities are responsible for adverse clinical events such as treatment non-responsiveness, but epigenetic modification might contribute a crucial role14. Emerging data suggest that microRNAs are one epigenetic mechanism that directly regulates KIT expression in GISTcells10. Limited patient data are available on the relationship between GISTs and miRNA expression in the Indian population. In the present study, we assessed the microRNA expression profiles of 50 GIST cases and investigated the relationships between these miRNAs and the molecular abnormalities and pathologic features of GISTs.
MiR-221 and miR-222 are two closely related microRNAs whose alteration has been observed in several cancers15. These miRNAs serve as both oncogenes and tumour suppressors depending on the tumour’s biology. It has been reported that the expression levels of miR-221 and miR-222 were significantly decreased in KIT-positive GISTs compared to normal tissue, whereas KIT-negative GISTs displayed the exact opposite expression profile16. Our data revealed that all four miRNAs were considerably down-regulated in GIST tumours relative to corresponding normal tissue. According to our findings, this inverse connection indicated that these miRNAs directly regulated KIT expression and may be key mediators of KIT in GISTs.
The link between KIT expression and the down-regulation of miR-221 and miR-222 in GIST supports the concept that these miRNAs suppressed the KIT receptor16. Numerous advanced cancers have been reported to have dysregulation of miR-221 and 222. MiR-221 and 222 up-regulation, as demonstrated in GIST, can prevent tumour growth in erythropoietic lineage cells by inhibiting KIT protein17. It has been reported that overexpression of miR-222 in GIST cell lines greatly inhibits cell proliferation, interrupts cell cycle progression, induces apoptosis and reduces overall KIT mRNA and protein levels18. Choi et al19 found a significant association between miR-494 expression and the KIT expression.
According to a study8, there was an inverse correlation between miR-494 and KIT expression in GIST. Further, it was observed that after transfection of GIST882 cells with miRNA mimics, overexpression of miR-494 significantly reduce KIT protein levels8. In addition, an in vitro investigation revealed that exogenous miR-494 induces KIT dysregulation more potently than miR-221 and miR-222. In contrast, the reduction of miR-494 induces KIT overexpression, indicating its potential role as a key regulator of KIT expression in GIST. Furthermore, our study identified distinct patterns of miRNA expression in different histological parameters. We found that each of these miRNAs was considerably downregulated in spindle subtypes, followed by epithelioid. MiR-221 and miR-222 were shown to be the most often dysregulated miRNAs in various GIST phenotypes. MiR-34a and miR-494 are additional essential microRNAs that regulate KIT/PDGFRA pathways. Moreover, all these miRNAs showed up-regulation in a few cases irrespective of primary tumour site, tumour size, c-KIT expression and other histological parameters. MiR-221, miR-222 and miR-494 were down-regulated in most gastric and small intestinal GIST. The expression profiling of these miRNAs did not reveal any relevant correlation with other histological features. It is important to investigate the correlation between the various molecular subtypes of GISTs and miRNA profiling. The present study examined the correlation with the KIT exons 9, 11, and 17and the PDGFRA exon 18. We observed significant associations between miRNA expression and KIT exons 11 and 17 mutant subtypes. On the other hand, miR-34a, predicted to target PDGFRA, was down-regulated in PDGFRA-mutated GIST. One of the most important facts concerning miRNA expression variations is that they have several target domains, making it challenging to determine a given miRNA’s specificity. Our results offer new insight into the association between miRNAs and histological features, enabling a more thorough understanding of GISTs at the molecular level.
The regulation of KIT or PDGFRA by microRNAs is of special interest. It can be concluded that miRNAs might regulate KIT expression by binding directly to the KIT mRNA4. Recently, numerous substantial functional investigations have been undertaken to validate the target genes of miRNA in GIST. Numerous studies have shown that essential genes involved in the pathophysiology of GIST, such as those in the KIT/AKT and PDGFRA pathways, are directly targeted by these miRNAs7. Thus miRNA dysregulation affects the expression of their target genes and regulates migration, proliferation and imatinib resistance via important biological processes10. It is important to emphasize that additional in vitro experiments utilizing GIST cell lines and the relevant molecular techniques are required to validate any target genes of miRNAs derived from this research. It might make sense to attribute this to the fact that various mechanisms, including changes in miRNA transcription or epigenetic modifications, can modify how miRNAs are expressed. As a result, the miRNA expression in particular cells or tissues varies, which in turn leads to protein and gene expression that is ideal for the corresponding cell state. Only the cell cycle protein called p27 has been demonstrated experimentally to be mediated by miR-221/222 in the cell lines of the prostate, papillary thyroid, and glioblastoma cancers16. Also, p57 has been experimentally identified as an additional target of miR-221 in hepatocellular cancer20. Based on all published evidence to date, the overexpression of miR-222, miR-221 and miR-494 in GIST has promising and successful therapeutic potential8,21. Many molecular anomalies are unique to GIST; however, only a few of these traits explain clinical behaviour and are significant for molecular classification.
Overall, our study demonstrated that all the four miRNAs were significantly down-regulated in GIST tumour compared to the corresponding normal tissue. These miRNAs showed significant correlation with c-KIT positive expression, spindle cell morphology, Gastric GIST, and KIT mutated GIST particularly exon 11. MiR-34a showed strong correlation with PDGFRA mutated GIST, while there was no significant correlation with other histopathological features. It is evident that these miRNAs might be targeting KIT and PDGFRA directly or indirectly. Our results offer new insight into the association between miRNAs and histological features, enabling a more thorough understanding of the classification and characterization of GISTs at the molecular level.
Financial support & sponsorship
None.
Conflicts of Interest
None.
Use of Artificial Intelligence (AI)-Assisted Technology for manuscript preparation
The authors confirm that there was no use of AI-assisted technology for assisting in the writing of the manuscript and no images were manipulated using AI.
References
- The genetic landscape of gastrointestinal stromal tumor lacking KIT and PDGFRA mutations. Endocrine. 2014;47:401-8.
- [CrossRef] [PubMed] [PubMed Central] [Google Scholar]
- Gastrointestinal tract spindle cell tumors with interstitial cells of Cajal: Prevalence excluding gastrointestinal stromal tumors. Oncol Lett. 2016;12:1287-92.
- [CrossRef] [PubMed] [PubMed Central] [Google Scholar]
- Gastrointestinal stromal tumors (GIST) related emergencies. Int J Surg. 2014;12:269-80.
- [CrossRef] [PubMed] [Google Scholar]
- MicroRNA involvement in gastrointestinal stromal tumor tumorigenesis. Curr Pharm Des. 2013;19:1227-35.
- [CrossRef] [PubMed] [Google Scholar]
- Understanding how miRNAs post-transcriptionally regulate gene expression. Prog Mol Subcell Biol. 2010;50:1-20.
- [CrossRef] [PubMed] [Google Scholar]
- Epigenetics in gastrointestinal stromal tumors: Clinical implications and potential therapeutic perspectives. Dig Dis Sci. 2013;58:3094-102.
- [CrossRef] [PubMed] [Google Scholar]
- miRNA-221 and miRNA-222 induce apoptosis via the KIT/AKT signalling pathway in gastrointestinal stromal tumours. Mol Oncol. 2015;9:1421-33.
- [CrossRef] [PubMed] [PubMed Central] [Google Scholar]
- MicroRNA-494 downregulates KIT and inhibits gastrointestinal stromal tumor cell proliferation. Clin Cancer Res. 2011;17:7584-94.
- [CrossRef] [PubMed] [Google Scholar]
- miRNA profiling in gastrointestinal stromal tumors: Implication as diagnostic and prognostic markers. Epigenomics. 2015;7:1033-49.
- [CrossRef] [PubMed] [Google Scholar]
- Small molecules in rare tumors: Emerging role of MicroRNAs in GIST. Int J Mol Sci. 2018;19:397.
- [CrossRef] [PubMed] [Google Scholar]
- A screen for epigenetically silenced microRNA genes in gastrointestinal stromal tumors. PLoS One. 2015;10:e0133754.
- [CrossRef] [PubMed] [PubMed Central] [Google Scholar]
- microRNA expression signatures of gastrointestinal stromal tumours: Associations with imatinib resistance and patient outcome. Br J Cancer. 2014;111:2091-102.
- [CrossRef] [PubMed] [PubMed Central] [Google Scholar]
- Analysis of relative gene expression data using real-time quantitative PCR and the 2- ΔΔCT method. Methods. 2001;25:402-8.
- [CrossRef] [PubMed] [Google Scholar]
- Interplay between epigenetics and metabolism in oncogenesis: Mechanisms and therapeutic approaches. Oncogene. 2017;36:3359-74.
- [CrossRef] [PubMed] [PubMed Central] [Google Scholar]
- Role of miR-221/222 in tumor development and the underlying mechanism. J Oncol. 2019;2019:7252013.
- [CrossRef] [PubMed] [Google Scholar]
- Down-regulation of miR-221 and miR-222 correlates with pronounced kit expression in gastrointestinal stromal tumors. Int J Oncol. 2011;38:503-11.
- [CrossRef] [PubMed] [Google Scholar]
- miR221/222 in cancer: Their role in tumor progression and response to therapy. Curr Mol Med. 2012;12:27-33.
- [CrossRef] [PubMed] [PubMed Central] [Google Scholar]
- Molecular mechanisms of imatinib resistance in gastrointestinal stromal tumor with focus on microRNAs [thesis]. Karolinska Institutet; 2015.
- MicroRNA expression profile of gastrointestinal stromal tumors is distinguished by 14q loss and anatomic site. Int J Cancer. 2010;126:1640-50.
- [CrossRef] [PubMed] [Google Scholar]
- miR-221-3p promotes hepatocellular carcinogenesis by downregulating O6-methylguanine-DNA methyl transferase. Cancer Biol Ther. 2020;21:915-26.
- [CrossRef] [PubMed] [PubMed Central] [Google Scholar]
- Non-coding RNAs, a novel paradigm for the management of gastrointestinal stromal tumors. Int J Mol Sci. 2020;21:6975.
- [CrossRef] [PubMed] [PubMed Central] [Google Scholar]




